
Sopot 1995
Wstęp
Przedmiot hydrobiologii
Hydrobiologia jest nauką o życiu w wodzie. Nazwa pochodzi od greckich słów: hydro - woda, bios - życie i logos - nauka. Zajmuje się ona organizmami żyjącymi w wodzie, ich wzajemnymi zależnościami i powiązaniem ze środowiskiem. Tak określony zakres tej dyscypliny stawia ją w kręgu nauk ekologicznych - nauk o współzależności organizmów i ich otoczenia. Wydzielenie hydrobiologii jako nauki spowodowane było specyficznymi cechami środowiska wodnego w porównaniu z lądowym, co ma swoje odbicie w biologii organizmów zamieszkujących to środowisko.
Hydrobiologia jest więc nauką kompleksową. Zadaniem jej jest określanie praw rządzących przebiegiem procesów w środowisku wodnym poprzez ustalanie wzajemnych zależności między organizmami wodnymi wynikających ze specyfiki ich budowy, trybu życia i rozmieszczenia a czynnikami ekologicznymi środowiska.
Czynniki te można rozpatrywać w czterech grupach:
· fizyczne (np. temperatura, światło, ruch wody, ciśnienie);
· chemiczne (np. gazy rozpuszczone w wodzie, elektrolity);
· edaficzne (np. właściwości podłoża i otoczenia zbiornika wodnego)
biotyczne (np. formy powiązań troficznych).
Hydrobiologia jest nowoczesną dziedziną wiedzy, dopiero od paru dziesiątków lat wykazującą intensywny rozwój. Wiele jest jeszcze problemów otwartych. Również niesprecyzowany jest do końca stosunek hydrobiologii do innych nauk przyrodniczych, co szczególnie jaskrawo widać w treści różnych podręczników poświęconych tej dyscyplinie wiedzy. Dawniejsi autorzy wiązali z hydrobiologią wszystkie wiadomości dotyczące życia roślin i zwierząt wodnych, łącznie z ich morfologią, fizjologią, czy systematyką. Nowsze podręczniki obejmują tylko te zagadnienia, które wykraczają poza szczegółowy zakres botaniki i zoologii a korzystają z fizyki, chemii oraz niektórych innych nauk - ostatnio także matematyki. W takim ujęciu hydrobiologia jest nauką niełatwą, gdyż do jej zgłębienia trzeba znać wiele innych dyscyplin mających charakter wyjściowy lub pomocniczy.
Do zrozumienia stosunków biotycznych panujących w hydrosferze konieczna jest znajomość samego środowiska, czyli wody jako ciała o swoistych właściwościach chemicznych i fizycznych, oraz poznanie indywidualnych cech hydrologicznych wód naturalnych. Tym zajmuje się hydrofizyka, hydrochemia, geologia, meteorologia i po części również hydromechanika. Wymienione wyżej dyscypliny naukowe, łącznie z takimi gałęziami nauk biologicznych jak botanika, zoologia, czy mikrobiologia, tworzą wspólną bazę rozwoju hydrobiologii.
Rozwój hydrobiologii
Wodą w swoim otoczeniu interesował się człowiek od najdawniejszych czasów. Jest ona zresztą podstawą rozwoju wszelkiego życia. Wszystkie cywilizacje rozwijały się nad wodami. Już w starożytności około 600 lat przed naszą erą panował pogląd, który głosił grecki filozof Tales z Miletu, jakoby woda była początkiem wszechrzeczy, tzn. wszystko na Ziemi "powstało z wody". Byłoby to zgodne z panującymi aktualnie teoriami o początkach życia na naszej planecie. Również z okresu starożytności pochodzą pierwsze opisy roślin i zwierząt wodnych zawarte w dziełach Arystotelesa (z ok. 322 roku p.n.e.) i Pliniusza Starszego (z 79 roku n.e.). Wykazy te znacznie poszerzyły się z chwilą wynalezienia w 1675 roku przez Antoniego van Leeuwenhoek mikroskopu, który umożliwił poznanie mikroskopijnych organizmów będących bardzo ważnym i istotnym składnikiem życia wód. Liczne opisy roślin i zwierząt wykonane przez mikroskopistów XVIII stulecia były punktem wyjścia dla pierwszych prac taksonomicznych, dotyczących m.in. glonów (wykonane przez Agardha), pierwotniaków (Ehrenberg, Dujardin, Stein), wrotków (Dutrochet, Hudson, Gosse) i skorupiaków (Müller, Schäffer, Sars). Badania te miały charakter wyłącznie morfologiczno-systematyczny i właściwie pomijały powiązania organizmów ze środowiskiem będącymi niezbędnym elementem współczesnej hydrobiologii.
Początki hydrobiologii datuje się dopiero na lata 70. ubiegłego stulecia. Wtedy to (1869 rok) profesor Uniwersytetu w Lozannie F.A.Forel rozpoczął studia nad fauną wodną Jeziora Genewskiego, uwzględniając jednocześnie w badaniach warunki fizyczno-chemiczne siedliska. Prace te zostały uwieńczone w latach 1892-1902 monografią tego jeziora, uważaną za pierwszy podręcznik hydrobiologii. W pracy tej po raz pierwszy użyto nazwy limnologia w odniesieniu do nauki o jeziorach. Forel wprowadził także do literatury nazwy trzech zasadniczych stref w jeziorze: przybrzeżnej - litoralnej, dennej - bentosowej i otwartej wody - pelagicznej i scharakteryzował warunki fizyczno-chemiczne panujące w każdej z nich. Podobne podejście cechowało prace znakomitego polskiego badacza Benedykta Dybowskiego. On to jako zesłaniec, w wyjątkowo trudnych warunkach, z dala od ośrodków naukowych, w latach 1873-1876 prowadził studia nad Bajkałem. Również w 1873 roku rozpoczął swoje badania nad jeziorem Mendota profesor Uniwersytetu stanowego Wisconsin E.A.Birge. Rok ten uznaje się za początek badań limnologicznych na kontynencie amerykańskim.
Do szybkiego rozwoju badań hydrobiologicznych przyczyniły się w zasadniczej mierze morskie wyprawy naukowe. Już podczas pierwszych ekspedycji wielkich odkrywców - Kolumba, Magellana, Cooka - zebrano wiele faktów dotyczących życia morskiego. Podczas słynnej podróży na statku "Beagle" Karol Darwin przeprowadził klasyczne obserwacje życia polipów koralowych i potem ogłosił swoją znaną teorię o powstawaniu raf koralowych. Jednakże dla rozwoju hydrobiologii podstawowe znaczenie miała słynna, trzyletnia wyprawa angielska (1873-1876) na statku badawczym "Challenger", w której uczestniczyli wybitni specjaliści z wielu dziedzin. Odegrała ona w procesie poznania życia w morzu rolę pionierską. Rok rozpoczęcia tej wyprawy uważa się z narodziny nowej dyscypliny wiedzy, jaką jest oceanologia - nauka o procesach przyrodniczych w morzu. Potem odbyło się wiele innych wypraw, organizowanych przez niektóre państwa, z programami już bardziej specjalistycznymi, dotyczącymi rozwiązania wąskich zagadnień. Taką była np. wyprawa na niemieckim statku "National" w 1889 roku, kierowana przez Viktora Hensena, zadaniem której było poznanie pożywienia ryb. Wykazano wtedy, że pokarmem dla ryb są drobne organizmy zwierzęce, które z kolei żywią się jeszcze mniejszymi organizmami roślinnymi. Te drobne, mikroskopijnej wielkości organizmy roślinne i zwierzęce Hensen w pracy ogłoszonej w 1887 roku nazwał planktonem. (patrz: rozdz. 6.1). Badacz ten opracował specjalną metodę łowienia planktonu za pomocą skonstruowanego przez siebie przyrządu, zwanego siecią planktonową. Również Hensen zaproponował rozwiązania metodyczne do obliczania ilości zawartych w morzu organizmów planktonowych stosowane do dziś. Wyniki badań Hensena stały się punktem wyjścia wielu prac zmierzających do określenia ogólnej produktywności morza.
Doniosłość poznawanych zagadnień, wcześniej dotyczących jedynie stosunków panujących w morzu, spowodowała rozszerzanie badań na akweny śródlądowe. I tak w 1896 roku Apstein wprowadził własną konstrukcję siatki do połowów planktonu słodkowodnego, zwaną dziś siatką Apstein’a. Tak szybki rozwój metod badawczych spowodował wyłonienie się nowej gałęzi hydrobiologii - nazwanej planktologią, czyli nauką o planktonie.
Hydrobiologia była w tych czasach zdominowana badaniami życia w toni wodnej. Spowodowało to słabszy rozwój innych jej kierunków. Przyczyna tego zastoju tkwiła w tym, że przez dłuższy czas nie umiano znaleźć odpowiedniej metody badania życia w innych środowiskach. Dopiero w 1910 roku duński badacz Petersen zastosował do połowu morskiej fauny dennej specjalny czerpacz dna, zamiast dotychczas stosowanych drag. W parę lat później szwedzki limnolog Ekman skonstruował analogiczny, tyle że mniejszy, czerpacz dna stosowany do zbierania makrofauny w zbiornikach słodkowodnych. Za pomocą tych przyrządów można wydobyć skrawek dna o określonej powierzchni, razem ze znajdującymi się tam organizmami. Można więc nie tylko określić skład gatunkowy, na co zresztą pozwalają zbiory z dragi, ale również określić liczebność poszczególnych gatunków.
Obserwacje hydrobiologów poczynione podczas ekspedycji miały charakter wycinkowy. Nie można było na ich podstawie badać zmienności zjawisk zachodzących w wodzie. Zaistniała więc konieczność prowadzenia systematycznych studiów obejmujących dłuższy okres, co stało się bezpośrednią przyczyną organizowania w różnych krajach stacjonarnych placówek naukowych położonych bezpośrednio nad morzem lub jeziorem, czy rzeką, tzw. stacji terenowych. Pierwsza stacja badawcza tego typu założona została już w 1872 roku w Neapolu przez Dohrna . Głównym jej zadaniem były badania morfologiczno-taksonomiczne i embriologiczne zwierząt zamieszkujących Zatokę Neapolitańską. W tym samym czasie rozpoczęła działalność także rosyjska stacja zoologiczna w Sewastopolu nad Morzem Czarnym.
Najstarsza stacja śródlądowa została założona przez Zachariasa nad jeziorem Plön na Pojezierzu Holsztyńskim w 1892 roku. Kierowana była przez wiele lat przez Thienemanna i stała się wówczas jednym z najpoważniejszych centrów badań hydrobiologicznych na świecie. Sławę zdobyła także prowadzona przez Ruttnera stacja w Lunz (Alpy austriackie). Po 1890 roku powstaje kilka nowych stacji hydrobiologicznych w Europie i Ameryce. Tuż przed drugą wojną światową było ich na świecie prawie 200. W Polsce o konieczności utworzenia stacji hydrobiologicznej wspominano już w 1888 roku. Jednak dopiero w 1920 roku powołano nowoczesną na ówczesne czasy Stację Hydrobiologiczną nad jeziorem Wigry na Suwalszczyźnie. Organizacyjnie była ona związana z Instytutem Biologii Doświadczalnej im. M. Nenckiego. Czynna była do roku 1939 i nie została reaktywowana po wojnie. Badania cenione na arenie międzynarodowej prowadzili w niej m.in.: A. Lityński, K. Demel, Z. Koźmiński, K. Passowicz, J. Wołoszyńska, J. Wiszniewski. Oprócz tego istniała założona w 1928 roku Stacja Biologiczna nad jeziorem Kiekrz koło Poznania, organizacyjnie związana z Uniwersytetem Poznańskim. Działał także rzeczny oddział Stacji Wigierskiej w Pińsku.
W okresie międzywojennym wzrosło w Polsce zainteresowanie problematyką morską. Przejawem tego było uruchomienie w 1923 roku Morskiego Laboratorium Rybackiego w Helu. Kierował nim przez wiele lat Kazimierz Demel - senior polskiej oceanografii. Były to pierwsze kroki polskiej nauki na Bałtyku. Badania zaczęto od podstawowych prac hydrograficznych i zwiadu rybackiego w wodach Zatoki Gdańskiej. W 1932 roku laboratorium przemianowano na Stację Morską. Spowodowane było to zmianą przynależności organizacyjnej, co okazało się bardzo dobrym pociągnięciem, które pozwoliło na rozszerzenie profilu badań o aspekty morsko-biologiczne i rybackie. Na początku 1939 roku stację przeniesiono do dużego i nowoczesnego gmachu w Gdyni. Po wojnie w tym właśnie budynku wznowiono działalność naukową pod nazwą Morskiego Laboratorium Rybackiego. W 1949 roku włączono je do Morskiego Instytutu Rybackiego, instytucji prężnie rozwijającej się, obejmującej zasięgiem swoich badań nie tylko cały Bałtyk, ale Morze Północne, Atlantyk i inne akweny, aż po wody Antarktydy i Pacyfiku.
Po drugiej wojnie światowej równie pomyślnie rozwijała się hydrobiologia na śródlądziu. Wynikało to z ogólnego trendu rozwoju nauki światowej i potrzeb gospodarki narodowej. Dziś hydrobiologia jako dyscyplina jest obecna prawie we wszystkich ośrodkach akademickich w Polsce, w instytutach Polskiej Akademii Nauk (m.in. w Instytucie Oceanologii w Sopocie) oraz w instytucjach resortowych, takich jak np. Instytut Rybactwa Śródlądowego w Olsztynie. Istotną datą dla rozwoju hydrobiologii wód morskich w Polsce było utworzenie kierunku studiów oceanograficznych na Uniwersytecie Gdańskim.
Środowisko wodne
Środowisko wodne tworzą zbiorniki wody naturalnej, czyli woda naturalna zebrana w określonej części geosfery. Na Ziemi wyróżnia się cztery geosfery o odmiennych właściwościach fizyczno-chemicznych. Są to:
· litosfera (skorupa ziemska),
· hydrosfera (płaszcz wodny),
· atmosfera (płaszcz gazowy otaczający Ziemię),
· biosfera (przestrzeń wokół Ziemi zasiedlona organizmami).
Zakresy logiczne trzech pierwszych pojęć wzajemnie się wykluczają, zaś czwartego obejmuje częściowo każde z nich i jest właściwie podstawą ich spójności. Zaznaczony na zielono na rys. 1. fragment hydrosfery jest polem zainteresowania hydrobiologów. Obejmuje on ogrom wód naturalnych, w których bytują organizmy żywe. Pod względem hydrologicznym wody te wykazują wielkie zróżnicowanie.
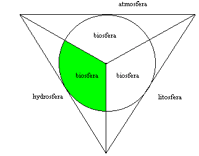
Rys. 1. Schemat analityczny zakresu logicznego geosfer
Woda naturalna występuje w przyrodzie w stanie ciekłym, stałym i gazowym. Stan ciekły to woda powierzchniowa zebrana w różnego typu zbiornikach wodnych, takich jak oceany, morza, jeziora i rzeki, oraz woda w glebie. Ta ostatnia może występować jako woda kapilarna, wypełniająca włosowate kanaliki między cząsteczkami gleby, lub jako woda wolna (grawitacyjna), będąca pod działaniem sił ciężkości, z której tworzy się woda gruntowa, zalegająca nad warstwami nieprzepuszczalnymi. Stan gazowy to para wodna, zaś stały stanowią lód i śnieg.
Zasadniczo można wyodrębnić wody morskie, zalegające obszary mórz i oceanów, i wody śródlądowe, wypełniające zbiorniki kontynentalne. Te ostatnie często określa się niesłusznie nazwą wód słodkich. Jest to nazwa archaiczna i uproszczona - znane są wcale nierzadkie przypadki, gdy wody kontynentalne są znacznie bardziej zasolone od wód oceanicznych.
Wody morskie obejmują obszar 95% wszystkich wód, a zalegają na powierzchni równej 2/3 powierzchni Ziemi. Stanowią one jeden wielki obszar wodny nazwany wszechoceanem. Obszar wszechoceanu wykazuje zróżnicowanie hydrograficzne. Jest ono mniej wyraziste niż wśród zbiorników kontynentalnych. Przeprowadzenie przejrzystego podziału wszechoceanu na akweny utrudnia brak logicznego kryterium, które doprowadziłoby do wyodrębnienia jasno różniących się kategorii zbiorników. Zasadniczo wyróżnia się trzy rejony morskie: oceany, morza właściwe i zatoki.
Oceany to najobszerniejsze rejony wszechoceanu o specyficznym systemie prądów, na ogół ograniczone zamkniętym układem kontynentów. Morza właściwe, stanowiące około 10% powierzchni wód, są to rejony wszechoceanu przylegające do kontynentów lub przez nie otoczone, zwykle płytsze od oceanów.
Morza właściwe dzieli się na kilka kategorii, w zależności od przyjętego kryterium. Ze względu na stopień związania z lądem wyróżnia się dwie grupy: morza przybrzeżne i śródziemne. Morza przybrzeżne przylegają do brzegu kontynentów, są oddzielone od oceanu podwodnymi progami lub szerokimi cieśninami (np. Morze Północne, Japońskie, Chińskie, Beringa). Morza śródziemne są otoczone lądem, a z wodami oceanicznymi połączone jedynie wąskimi i zwykle płytkimi cieśninami (np. Bałtyk, Morze Śródziemne, Morze Czarne). W grupie mórz śródziemnych można wyodrębnić morza międzykontynentalne, otoczone lądem odrębnych kontynentów, zwykle głębokie (np. Morze Śródziemne, Morze Czarne, Morze Czerwone), i morza wewnątrzkontynentalne, otoczone lądem jednego kontynentu, zwykle płytkie (np. Bałtyk, Morze Białe, Zatoka Hudsona). Ze względu na głębokość wyróżnia się morza płytkie (szelfowe), wykształcone na platformie kontynentalnej, czyli szelfie, o średniej głębokości 200 m (np. Bałtyk, Morze Północne), oraz morza głębokie, obejmujące także rejony głębsze poza platformą kontynentalną (np. Morze Śródziemne). Stosuje się jeszcze inne kryteria podziału, np. położenie geograficznie (morza gorące, umiarkowane i zimne).
Zatoki są akwenami, które nie dają się ściśle zdefiniować. Niektóre z nich, np. Zatoka Biskajska, Zatoka Hudsona, czy Zatoka Bengalska geograficznie mogą być uważane za morza przybrzeżne. Inne zaś mają charakter rzeczywiście bardziej zatokowy. Są to małe, daleko wrzynające się w ląd części mórz.
Równie trudna jest klasyfikacja zbiorników śródlądowych, które często łączą się i przechodzą jedne w drugie. Przyjął się ogólny podział na wody stojące (lenityczne) i płynące (lotyczne). W każdej z tych dwóch kategorii wyróżnia się kilka typów zbiorników. Do pierwszej zalicza się zbiorniki o różnej wielkości, ale z wodą stagnującą. Są nimi nie tylko jeziora lub stawy, w których woda występuje stale, ale również kałuże zawierające wodę zazwyczaj tylko okresowo, a także wody podziemne, zgromadzone w gruntowych warstwach wodonośnych, w jaskiniach lub szczelinach skał. Do drugiej kategorii zalicza się wszystkie cieki, takie jak: źródła, potoki, strumienie i rzeki, kończące się zwykle ujściem do morza, w którym wykształca się specyficzny obszar wód słonawych, wskutek tzw. cofki wód morskich. Stwarza to organizmom odmienne niż w rzece warunki bytowania. Do tej kategorii zalicza się również cieki sztuczne, zbudowane przez człowieka - kanały, rowy melioracyjne oraz zbiorniki zaporowe, które wiążą w sobie wiele cech jezior i cieków.
Hydrosfera stanowi ponad 70% powierzchni Ziemi. Całkowitą objętość wody na Ziemi ocenia się następująco:
· oceany i morza – 1370 x 106 km3
· lodowce – 20 x 106 km3
· woda w litosferze – 0,5 x 106 km3
· woda w atmosferze – 13 x 103 km3
Woda ta jest w ciągłym ruchu ulegając wymianie między stanem ciekłym, stałym i gazowym wskutek oddziaływania energii cieplnej Słońca oraz sił ciężkości. Krążenie wody w przyrodzie opisuje cykl hydrologiczny (rys. 2). Z jego analizy wynika, że ogólna ilość wody w przyrodzie jest stała - jest to cykl zamknięty. Pod wpływem promieniowania słonecznego woda z powierzchni mórz i oceanów oraz z kontynentów paruje (E), dostając się do atmosfery. Skondensowana w postaci chmur powraca jako opad (P). Tak więc cykl hydrologiczny można wyrazić w postaci bilansu wodnego, gdzie całkowite parowanie równe jest całkowitemu opadowi atmosferycznemu, a więc E=P.

Rys. 2. Cykl hydrologiczny
W cyklu hydrologicznym krąży zaledwie 577 tys. km3, co stanowi niecałe 0,05% ogólnych zasobów wodnych hydrosfery. Pozostała ilość wody jest zatrzymywana (retencjonowana) na powierzchni i okresowo wyłączona z obiegu. Z ogólnej puli wody krążącej 80% bierze udział w obiegu między oceanem i atmosferą, a 20% między lądem a atmosferą. Tak więc bilans wodny fazy oceanicznej można przedstawić równaniem 1
Równanie 1.
![]()
zaś fazy lądowej 2
Równanie 2.
![]()
gdzie:
PO - opad na powierzchnię oceanów
PK - opad na obszary lądów
EO - parowanie z powierzchni oceanów
EK - parowanie z powierzchni lądów
HK - odpływ z lądów do oceanu światowego
ΔRK - zmiany retencji wody na lądach
ΔRO - zmiany retencji wody w oceanie.
W odpływie wody z lądów wyodrębnia się odpływ powierzchniowy z dorzeczy do mórz oraz odpływ podziemny z wód gruntowych. Wskutek odpływu wody z kontynentów do oceanu światowego faza lądowa cyklu hydrologicznego charakteryzuje się przewagą opadu atmosferycznego nad parowaniem, natomiast w fazie oceanicznej stosunki układają się odwrotnie.
Tempo wymiany wód w cyklu hydrologicznym jest różne i zależy od rodzaju wód naturalnych. Uważa się, że teoretycznie woda w hydrosferze jako całości wymienia się przeciętnie co 2800 lat. W oceanach całkowita wymiana wody szacowana jest na 3-4 tys. lat. Przyjmuje się, że na lądzie całkowita wymiana zasobów wilgoci glebowej następuje co roku. Okres wymiany wód jeziornych trwa ok. 3. lat, rzecznych od 12 do 25 dni. Najdłuższym okresem całkowitej wymiany, sięgającym 16.000 lat charakteryzują się lodowce i wieczne śniegi, zaś największą aktywnością odznacza się woda atmosferyczna, a ściślej para wodna w atmosferze. Ulega ona wymianie co 8-10 dni.
Zasoby wody na lądach nie są rozłożone równomiernie. Jej niedobory w niektórych rejonach Ziemi wynikają ze specyfiki warunków klimatycznych. Miara zasobów wodnych może być ilość wody przypadającej rocznie na jednego mieszkańca. Wynosi ona w Polsce około 1.700 metrów sześciennych, podczas gdy we Francji 3.000, w Niemczech 2.500, a w Rosji prawie 7.000. Tak więc w porównaniu z krajami sąsiednimi sytuacja w Polsce jest krytyczna. Wynika on z faktu, iż Polska leży w strefie pomiędzy klimatem morskim (atlantyckim) a kontynentalnym. Są obszary, szczególnie centralna część Polski, gdzie bilans wodny jest dalece niezrównoważony.
Właściwości fizyczne wody
Budowa cząsteczki wody
Pod względem właściwości fizycznych woda wykazuje anomalia. W przeciwieństwie do wszystkich innych płynów, podczas zamarzania nie kurczy się, a powiększa objętość. Ciężar wody chemicznie czystej jest największy nie przy 0oC, lecz przy 4oC. Przez wiele lat nie umiano sobie wytłumaczyć tych zjawisk. Dopiero dzięki rozwojowi chemii fizycznej odkryto specyficzną budowę molekuły wody. Budowa cząsteczki wody pomimo, że na pozór prosta (H20), jest bardzo złożona i wciąż niedostatecznie rozpoznana.
Zasadniczą cechą budowy molekuły wody jest niesymetryczne rozmieszczenie atomów wodoru wokół tlenu. Atomy te znajdują się pod kątem 105o w stosunku do atomu tlenu. Powoduje to asymetrię w rozmieszczeniu ładunków elektrycznych i wytwarza cząsteczki o dwóch różnie naładowanych biegunach, tzw. dipol (rys. 3). Wokół cząsteczki wody powstaje pole elektryczne. Dzięki niemu następuje łączenie się cząsteczek wody przeciwnymi biegunami, a także wiązanie ze spolaryzowanymi cząstkami innych substancji. Mogą więc tworzyć się układy większych skupień wokół molekuł wody. Wiadomo, że molekuła wody o wzorze H20 istnieje tylko w stanie gazowym - jako para wodna. Natomiast w stanie ciekłym i stałym istnieją drobiny o sumarycznym wzorze H4O2, H6O3 itd. Siłę przyciągania przez dipol wody innych substancji nazywa się momentem dipolowy wody. Jego wartość stanowi iloczyn odległości między elektrycznymi środkami ciężkości ładunków dodatnich i ujemnych w cząsteczce i wielkości ładunku.
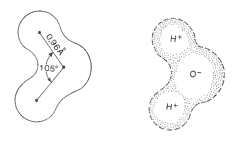
Rys. 3. Budowa cząsteczki wody
Inną właściwością budowy cząsteczek wody, związaną z możliwością ich skupiania się, jest zdolność do tworzenia się wiązań wodorowych (rys. 4). Wynika to ze skomasowania w protonie wodoru na małej powierzchni dużego ładunku, który odkształca osłonę elektronową silnie elektroujemnego tlenu i wiąże się z nim elektrostatycznie. Tak więc atom wodoru, nie tracąc połączenia z tlenem, do którego przynależy, tworzy połączenia, w których każdy atom tlenu koordynuje tetraedrycznie cztery atomy wodoru.
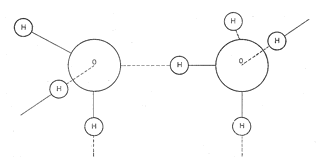
Rys. 4. Wiązania wodorowe łączące molekuły wody.
Większość nowoczesnych teorii zakłada, że skupienia molekuł wody mogą występować w trzech postaciach:
- ułożenie ich przypomina postać krystaliczną kwarcu (SiO4), tzw. trihydrol, forma głównie występująca w lodzie;
- postać krystaliczna jak w krzemionce (SiO2), czyli tzw. dihydrol.
- skupienia molekuł tworzących nitkę, tzw. monohydrol.
Zawartość w wodzie wielocząsteczkowych agregacji zmniejsza się wraz ze wzrostem temperatury. W wyższych temperaturach wody prędkość ruchu cząsteczek wzrasta i siły przyciągania nie są w stanie utrzymać ich obok siebie. Trihydrol występuje w temperaturze poniżej 4oC, jednak nawet w tej temperaturze dihydrol stanowi 50% składu. Wraz ze wzrostem temperatury zawartość trihydrolu maleje - do 30% w 4oC i 5% w temperaturze 15oC. W przypadku monohydrolu sytuacja jest odwrotna, w temperaturze 15oC stanowi zaledwie 15% i jego zawartość szybko zwiększa się wraz ze wzrostem temperatury, osiągając prawie 100% w punkcie wrzenia.
W wodzie naturalnej najliczniejsze są zwykłe cząsteczki wody o ciężarze cząsteczkowym 18 (O16, H1, H1). Stanowią one 99,8%. Pozostały nikły odsetek stanowią cząsteczki, w których skład wchodzą izotopy tlenu o ciężarze 17 i 18 oraz wodoru o ciężarze 2 i 3. Zwiększenie liczby cząsteczek cięższych powoduje zwiększenie się gęstości wody.
Ruch wody
Ruch mas wodnych jest ważnym czynnikiem ekologicznym wpływającym szczególnie wyraźnie na rozmieszczenie organizmów w wodzie oraz na kształtowanie geomorfologiczne zbiorników wodnych. Woda jako środowisko 775 razy cięższe od powietrza wywiera silne działanie mechaniczne na przedmioty znajdujące się na drodze jej ruchu. Zjawisko przemieszczania się mas wodnych jest wieloaspektowe i ma zróżnicowane przyczyny. Jest ono kierunkowe i stąd określa się je nazwą prądu.
W jeziorach prądy tworzą się głównie pod wpływem wiatrów, rzadziej jako wynik różnic temperatur lub następstwo kolebania się (tzw. sejsze). Wiatr powoduje w jeziorach wytwarzanie się prądów falowych i prądów cyrkulacyjnych (patrz rozdz. 3.4). Siła falowania zależy od stopnia oddziaływania wiatru, która jest funkcją siły wiatru, wielkości powierzchni jeziora, stopnia osłonięcia brzegu itp. Jeziora rozległe i płytkie podlegają silnemu falowaniu. Fale docierające do brzegu w zależności od siły prądu niszczą go tworząc kipiel, czyli prądy wirujące. Woda w ten sposób odbita od brzegu rozchodzi się na boki lub wnika w głąb, dając prąd cyrkulacyjny. Wywołuje to przenoszenie materiału skalnego erodowanego w jednym miejscu na stronę nawietrzną, gdzie jest on osadzany. Proces taki powoduje wypłycanie brzegów i tworzenie się ławicy przybrzeżnej. Obok prądów poziomych występują prądy pionowe, konwekcyjne, czyli wyrównawcze. Ich istnienie jest związane z różnicami gęstości wody w różnej temperaturze.
Zjawisko określane jako sejsze lub kolebanie się wody występuje pod wpływem działania gwałtownych zmian ciśnienia atmosferycznego, silnych deszczy lub wiatrów. Wówczas część powierzchni jeziora się obniża, co przy bezwładności całej masy wodnej dążącej do wyrównania w poziomie powoduje równocześnie podniesienie się poziomu w innym rejonie. Po ustąpieniu przyczyny następuje stopniowe wyrównywanie się poziomu czemu towarzyszy kolebanie się mas wodnych, a w efekcie powstanie fali stojącej, mającej jeden lub kilka węzłów (rys. 5).
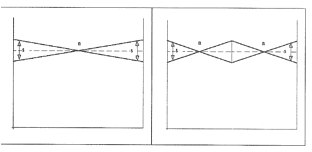
Rys. 5. Sejsze: fala jednowęzłowa (z lewej); fala dwuwęzłowa (z prawej)
W rzekach mamy do czynienia z prądem mechanicznym, grawitacyjnym, który powstaje w wyniku grawitacyjnego spływu masy wód z miejsc wyżej położonych w dół. Prąd wody w rzece jest hamowany tarciem o podłoże i stąd prędkość jego jest nierówna. Najsilniejszy jest w górnej warstwie środkowej części nurtu rzeki, a najsłabszy przy dnie i brzegach. W miejscach przeszkód (duże kamienie lub powalone drzewa) tworzą się pionowe i boczne prądy wirowe, powodujące przenoszenie materiału dennego. Dno ruchliwe i erodowane brzegi rzeki są niekorzystnym biotopem do zasiedlania.
W morzach występują trzy zasadnicze typy ruchu mas wodnych. Są to: pływy, falowanie i prądy wewnętrzne. Pływy są pionowymi ruchami wody morskiej, o ściśle określonym i okresowym charakterze, wywoływanymi siłami kosmicznymi tj. działaniem grawitacyjnym Księżyca i Słońca. W punkcie Ziemi, gdzie odległość do Księżyca jest najmniejsza, a więc jego oddziaływanie największe, następuje wypuklenie mas wodnych, spowodowane przesunięciem wody z innych miejsc (rys. 6). Pływy są więc swego rodzaju falą wzniesionej wody, przechodzącą wokół Ziemi z częstotliwością obrotu Księżyca. Wskutek obrotowego ruchu Ziemi i Księżyca wokół ich wspólnego środka masy powstaje drugie symetryczne podniesienie wód w miejscu najbardziej odległym od Księżyca. Sprawia to, że przez każdy punkt na Ziemi przechodzą dwie fale wysokiej wody, czyli przypływu w odstępie 12 godz. i 25 min. W punktach położonych prostopadle do przypływu następuje obniżenie poziomu wody, czyli odpływ. Zjawiska przypływu i odpływu są wyraźnie widoczne na brzegach oceanów i mórz otwartych. Im akwen jest bardziej zamknięty, śródlądowy, tym zjawisko to ma mniejszy zasięg, np. w Bałtyku jest ono minimalne i nie ma ono znaczenia ekologicznego.
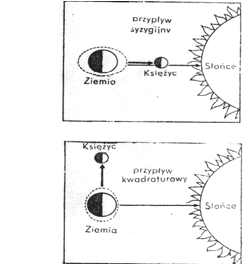
Rys. 6. Zjawisko pływów. Linia przerywana wyobraża poziom wody.
Wysokość fali pływowej uzależniona jest od wzajemnego ustawienia Ziemi, Księżyca i Słońca. Pomimo swej ogromnej masy Słońce wywiera znacznie mniejszy wpływ na wodę. Wynika to z dużej odległości dzielącej Słońce od Ziemi. Gdy Ziemia, Księżyc i Słońce znajdują się w jednej linii, czyli podczas nowiu lub pełni w ułożeniu zwanym syzygią, siły przyciągające Księżyca i Słońca kumulują się i wywołują wielkie pływy syzygijne (rys. 7). W okresach kwadratury, t.j. pierwszej i ostatniej kwadry, następuje osłabienie się wzajemne sił Księżyca i Słońca. Mamy wówczas do czynienia małymi pływami kwadraturowymi.
Rejon pływów jest specyficznym biotopem. Mogą w nim bytować organizmy, które w wyniku ewolucji zaadaptowały się do warunków okresowego odsłaniania. Muszą więc one znajdować schronienie np.: w wilgotnym piasku, kępach glonów lub drobnych zastoiskach wody na nierównym dnie. Trzeba pamiętać, że w wyniku parowania wody w tych minisiedliskach następuje dość znaczny wzrost zasolenia - przeżyć w takich warunkach mogą tylko organizmy mające zdolność przystosowania się do zmiennego zasolenia.
Falowanie powierzchniowe spowodowane wiatrem ma różny stopień nasilenia. Zależy od siły wiatru. Falowanie z dala od brzegów nie wywołuje przesuwania się cząsteczek wody w poziomie, lecz jedynie oscylujący pionowo ruch kolisty. Tylko przy brzegu fale wstrzymywane przez ląd spiętrzają się, tworząc przybój i mechanicznie odziałując na brzeg. Niszczycielski charakter falowania jest szczególnie wyraźny podczas sztormu.
Prądy morskie można porównać do olbrzymich rzek płynących określonymi "korytami" w bezkresie spokojnych wód wszechoceanu. Wśród przyczyn powstawania prądów wymienia się: jednokierunkowy obrót Ziemi dookoła osi, wiatry wiejące z określonych kierunków, różnice gęstości wody spowodowane ciepłem słonecznym lub lodami polarnymi. System prądów morskich tworzy skomplikowaną cyrkulację w każdym oceanie. Obejmuje on prądy powierzchniowe powstające pod wpływem wiatru, oraz prądy głębinowe o innym pochodzeniu. Prądy morskie mają zasadniczo przebieg poziomy, chociaż istnieje także cyrkulacja pionowa. Typowym jej przykładem jest upwelling, czyli inaczej prąd wstępujący. Jest to proces, w którym zimna woda z głębi jest wynoszona na powierzchnię. Dzieje się to np. w rejonach przybrzeżnych stref podzwrotnikowych i jest efektem wiania jednokierunkowych, odlądowych wiatrów. Znaczenie biologiczne tego zjawiska jest ogromne. W jego wyniku wody wgłębne, bogate w sole odżywcze wydostają się na powierzchnię, gdzie zachodzi proces produkcji materii organicznej. Stąd też są to jedne z najżyźniejszych rejonów wszechoceanu.
Warunki świetlne
Światło, a właściwie energia promienista, ma podstawowe znaczenie w procesie produkcji organicznej w wodzie. Energia ta pochodzi oczywiście z promieniowania słonecznego, które na powierzchni Ziemi składa się z promieniowania bezpośredniego i rozproszonego. To pierwsze uzależnione jest od kilku czynników, takich jak szerokość geograficzna, pora dnia, przezroczystość atmosfery. Promienie padające na powierzchnię wody mogą do niej przenikać, zostać odbite od powierzchni lub być rozproszone. Stopień odbicia promieni zależy od kąta ich padania na powierzchnię wody (rys. 7). Woda odbija więcej promieni w godzinach rannych i wieczornych, gdy padają one bardziej ukośnie, niż w południe.
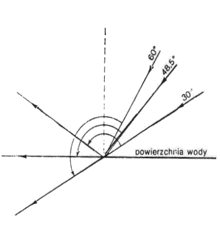
Rys. 7. Przenikanie i odbijanie od wody promieni świetlnych w zależności od kąta padania
Ilość odbijanego od powierzchni wody światła zależy także od pory roku. W naszej szerokości geograficznej latem od powierzchni wody odbija się około 2,5-3% promieni, zimą zaś prawie 14%. Dane te odnoszą się do powierzchni gładkiej - sfalowanie powierzchni wody proces ten znacznie komplikuje. Jeszcze trudniejsze jest określenie ilości promieniowania rozproszonego dochodzącego do powierzchni wody. W zależności od stopnia zachmurzenia może ono stanowić 8-100% światła padającego na zbiornik. Długość fali promieni widzialnych nie wpływa na stopień odbicia.
Promienie nie odbite przenikają do zbiornika wodnego, ponieważ woda jest ciałem przezroczystym. W wodzie następuje ich pochłanianie. Promienie światła białego są w wodzie pochłaniane selektywnie. Najszybciej pochłaniane są promienie fali długiej - podczerwone i czerwone, oraz fali krótkiej - fioletowe i ultrafioletowe. Głębiej przenikają promienie pomarańczowe i niebieskie, a najgłębiej żółto-zielone. Tak więc zasięg światła maleje wraz z głębokością wody (rys. 8).
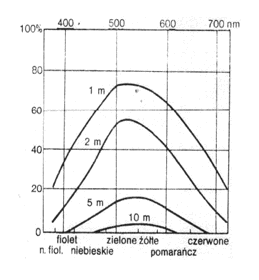
Rys. 8. Intensywność pochłaniania różnych części widma słonecznego w wodzie jeziornej
Proces przenikania promieni zależy od rodzaju i ilości soli rozpuszczonych w wodzie oraz obecności zawiesiny. Takie związki chemiczne jak chlorki wapnia i magnezu zmniejszają pochłanianie promieni. Dlatego też wody morskie bogate w te sole są bardziej przejrzyste od innych. Obecność amoniaku i azotanów zmniejsza przepuszczalność wody dla promieni.
Poznanie zjawiska przenikania promieniowania uzależnione było od dokładności metod pomiarowych. Najprościej zasięg światła można określić za pomocą krążka Secchiego. Jest to biały krążek metalowy o średnicy 20 cm spuszczany na linie. Głębokość, na której przestaje być widoczny, pomnożona przez 2 daje informacją o zasięgu promieniowania widzialnego. Nowsze metody, stosowane szczególnie w badaniach morskich, polegają na użyciu papieru fotograficznego umieszczonego w odpowiednich kamerach. Metody te są szczególnie przydatne przy pomiarach bardzo słabego natężenia światła. W najnowocześniejszych metodach stosuje się fotokomórki.
Przenikanie promieni świetlnych zależy także od barwy i przezroczystości wody, czyli od ilości zawiesiny i związków koloidalnych, a zwłaszcza humusowych, barwiących wodę na brunatno. Od rzeczywistej barwy wody, czyli barwy własnej trzeba odróżnić barwę pozorną, pochodzącą z odbijania się w lustrze wody nieba i chmur oraz okolicznych skał lub lasów. Barwa własna wody jest efektem stopnia rozproszenia światła. Najsilniej rozpraszane są promienie niebieskie, najsłabiej czerwone. Stąd też woda wolna od zanieczyszczeń ma barwę zielononiebieską lub, gdy jest bardzo czysta, nawet szafirową. Wówczas właśnie te promienie jako najbardziej rozproszone docierają do oka obserwatora. Taką barwą charakteryzują się wody czystych jezior górskich. Na nizinach wody zawierają stale wiele rozpuszczonych związków organicznych - nie mają więc takiej barwy. Ilość związków organicznych stanowi o czystości wody, stąd też znajomość barwy ma znaczenie praktyczne. Forel wprowadził, a Ule później rozszerzył, skalę pozwalającą na ocenę barwy wody. Skala Forela-Ulego obejmuje razem 21 barw. Woda jezior górskich i otwartych mórz odpowiada numerom I-V. Bardzo czyste jeziora niżowe mają wodę z zakresu VI-IX. Większość jezior europejskich ma wodę o różnych odcieniach barwy żółtej, co mieści się w skali numerów X-XIV. Większe numery skali obejmują zbiorniki o wodzie mniej lub bardziej brunatnej.
Przezroczystość wód naturalnych jest bardzo zmienna. Zależy ona od opadów atmosferycznych, znoszonych z lądu cząstek gleby, pyłów nawianych wiatrem, wpływających do zbiornika ścieków, zakwitów planktonowych itp. Wszystko to razem tworzy zawiesinę. Im jest ona gęstsza, tym wygaszanie światła następuje szybciej. Utrzymanie się zawiesiny w wodzie, zgodnie z prawem Stoksa, uzależnione jest od wielkości cząstek i różnicy ich ciężaru właściwego w stosunku do ciężaru właściwego wody. Tak więc cząstki mineralne, takie jak ziarna piasku, opadają szybciej niż takich samych rozmiarów cząstki obumarłych organizmów. Natomiast wiele organizmów stanowiących tzw. "żywą zawiesinę" wyłamuje się spod tego prawa i dzięki zdolności samodzielnego ruchu utrzymuje się na danym poziomie lub może podpływać w górę. Również ciała koloidalne, silnie uwodnione, utrzymują się długo w zawiesinie.
Uwzględniając całość stosunków świetlnych można wydzielić w morzach i głębokich zbiornikach jeziornych trzy warstwy poziome. Warstwę górną, dobrze naświetloną, o dobrych warunkach do fotosyntezy, zwaną eufotyczną. Głębiej wykształca się warstwa pośrednia, o nikłych ilościach światła, zwana dysfotyczną lub inaczej warstwą cienia albo półmroku. Niżej zalega rozległa warstwa mroku, całkowicie pozbawiona światła, zwana afotyczną. Dla oceanów można przyjąć, iż warstwa eufotyczna sięga średnio do głebokości 50-80 m, dysfotyczna do 350-400 m, zaś głębiej rozciąga się obszerna warstwa afotyczna. Miąższość strefy eufotycznej w morzach jest mniejsza, szczególnie w strefach przybrzeżnych, gdzie więcej jest zawiesiny w wodzie. Również w jeziorach wartości te są inne. W jeziorach niżowych strefy umiarkowanej warstwa eufotyczna nie przekracza zwykle głębokości 5-10 m. W wielu zbiornikach bywa jeszcze płytsza i może nie dochodzić do głębokości 1 m. Także w rzekach, ze względu na dużą ilość zawiesiny transportowanej wodami rzek, warstwa ta jest bardzo płytka.
Stosunki świetlne w zbiorniku determinują procesy produkcyjne. Ogólne wykorzystanie światła do tych celów jest małe i wynosi zaledwie ok. 2%. Fotosynteza najkorzystniej przebiega w warstwach nieco głębszych, na głębokości 0,5-2 m zależnie od stopnia przezroczystości wody. Część promieniowania świetlnego - ultrafiolet i podczerwień - jest szkodliwa zarówno dla roślin jak i zwierząt. W wodzie czystej te długości fali świetlnej są prawie całkowicie wygaszane już na głębokości 1 m. Większemu ograniczeniu tego szkodliwego wpływu światła sprzyja zawiesina i wapń rozpuszczony w wodzie. W przypadku dużej ilości zawiesin promienie ultrafioletowe i podczerwone mogą być pochłonięte w całości już na głębokości 10 cm. Czynnikiem uodparniającym organizmy zwierzęce na działanie tych promieni jest wapń - w wodach ubogich w ten pierwiastek plankton jest zazwyczaj słabiej rozwinięty. Tak jak nadmiar światła, również jego brak działa niekorzystnie. W wodach silnie zacienionych produkcja biologiczna bywa mała. Obniżenie produkcji mogą powodować same organizmy żywe zasłaniając strumień światła dopływający do wody. Masowy rozwój glonów planktonowych tworzących tzw. zakwity, w zależności od stopnia intensywności, może znacznie osłabić penetrację światła i ograniczyć przez to produkcję. Podobnie rośliny naczyniowe o liściach pływających (np. grążele), czy niezakorzenione (np. rzęsy), mogą całkowicie odciąć dopływ światła do zbiornika wodnego.
Do najbardziej powszechnych zjawisk w zbiornikach wodnych należą wędrówki pionowe zwierząt planktonowych. Są one zsynchronizowane z cyklem dobowym dnia i nocy. W warstwach powierzchniowych ilość planktonu zwierzęcego wyraźnie zmniejsza się w godzinach południowych, a zwiększa wieczorem lub nocą. Zasięg wędrówki jest różny - od płytkich 40-metrowych wędrówek w jeziorach, do głębokich wykonywanych np. przez pospolitego widłonoga morskiego Calanus finmarchicus, który opuszcza się w ciągu dnia do głębokości 400-500 m, a pod wieczór podnosi się znów pod powierzchnię (rys. 9). Jednak nie wszystkie zwierzęta planktonowe wykonują takie wędrówki, a jedynie te, które wykazują fototaksję ujemną. Dlatego też organizmy można podzielić na fotofile i fotofoby, czyli formy zdążające za światłem lub go unikające. Ponadto wyróżnia się dość znaczną grupę zwierząt euryfotycznych, znoszących bez szkody szeroki zakres wahań natężenia światła.
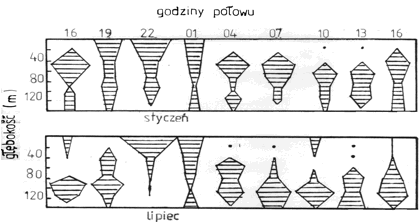
Rys. 9. Przebieg wędrówek dobowych zooplanktonu na przykładzie samic u Calanus finmarchicus
Innym przykładem przystosowania się organizmów do specyficznych warunków świetlnych wód naturalnych jest występowanie na styku warstwy eufotycznej z dysfotyczną, czyli w strefie światła zielono-niebieskiego, glonów o chromatoforach żółtawych, brunatnych lub czerwonych. Tak więc mają one zabarwienie dopełniające do barwy światła tam docierającego. Zjawisko to określa się przystosowaniem lub adaptacją chromatyczną. Również organizmy zwierzęce bytujące w warstwie dysfotycznej wykazują przystosowania anatomiczne narządu wzroku, wynikające z ograniczonej ilości występującego tam światła. Przystosowania te idą w dwóch kierunkach: wydłużenia osi optycznej i wykształcenia dużej soczewki. Są więc to tzw. oczy teleskopowe, dzięki którym zwierzę rozpoznaje dokładnie przedmioty z bliska. Przedmioty odległe, wskutek panującego mroku, i tak są niedostrzegalne.
Innym zjawiskiem świetlnym, nie mającym co prawda nic wspólnego z promieniowaniem słonecznym, jest powszechna zwłaszcza w środowisku morskim bioluminescencja. Wiele organizmów bytujących w warstwach głębinowych posiada specjalne narządy świecące o zdumiewającym bogactwie kształtów. Stwierdzono, że ponad 10% ryb głębinowych ma zdolność świecenia. Również wiele występujących w planktonie organizmów jednokomórkowych wykazuje zjawisko bioluminescencji. Do nich należą przede wszystkim bruzdnice oraz wiele innych wiciowców, a także żebropławy, meduzy i skorupiaki. W morzach strefy gorącej zjawisko bioluminescencji mogą wywołać masowo występujące bakterie, które znaleziono również w ciele świecących osłonic i głowonogów. Istnieją poglądy wiążące zjawisko bioluminescencji z obecnością bakterii w ciele organizmu świecącego. Bakterie te mieszczą się w fotoforach, gdzie w wyniku procesu chemicznego, w którym bierze udział specjalny związek lucyferyna i enzym lucyferaza, wytwarza się to "zimne światło". Na temat ekologicznego znaczenia tego zjawiska istnieje wiele hipotez, niekiedy wzajemnie sprzecznych.
Warunki termiczne
Temperatura jest jednym z podstawowych czynników ekologicznych w wodzie. W zależności od temperatury zmieniają się takie parametry fizyczne, jak gęstość i lepkość wody, które istotnie wpływają na bytujące w niej organizmy. Znacznym zmianom podlega gęstość (masa właściwa) wody. W temperaturze ujemnej przy normalnym ciśnieniu atmosferycznym i zasoleniu równym 0, woda w postaci lodu ma gęstość zaledwie 0,9167 g/cm3, a wartość 1,00 osiąga w temperaturze +4oC. Oznacza to, że woda marznąc zwiększa swoją objętość. W temperaturze powyżej +4oC również następuje spadek gęstości wody, ale dopiero w temperaturze +30oC osiąga ona gęstość lodu (rys. 10). Zasada ta odnosi się do wody destylowanej. W wodzie naturalnej, w której są rozpuszczone różne substancje, wygląda to nieco inaczej. Drugi z omawianych parametrów fizycznych, lepkość zmniejsza się przy wzroście temperatury; podniesienie się temperatury o 10oC zmniejsza ją przeciętnie o połowę. Ma to duże znaczenie dla unoszenia się i pływania organizmów planktonowych w toni wodnej. Tak więc, gdyby nie inne czynniki, to w temperaturze +20oC organizmy opadałyby dwa razy szybciej niż w temperaturze 0oC, .
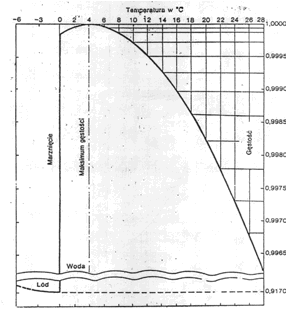
Rys. 10. Zmienność gęstości wody czystej w zależności od temperatury
Na procesy życia organizmów wodnych wpływa nie tylko temperatura, ale także ilość ciepła zawartego w wodzie. Każdy proces biologiczny, np. dojrzewanie jaj, rozwój zarodków i wzrost, wymaga określonej ilości ciepła. Nagłe zmiany temperatury powodują jego zachwianie, prowadząc nawet do śmierci. Na szczęście jednak woda charakteryzuje się dużym bezwładem cieplnym, tzn. ogrzewa się bardzo powoli i tak samo wolno oddaje ciepło. Ta duża pojemność cieplna łagodzi szybkość zmian temperatury. Zmiany te zachodzą stopniowo, co ułatwia organizmom przystosowanie się do nowych warunków termicznych.
Źródłem ciepła w zbiorniku wodnym jest promieniowanie słoneczne padające na jego powierzchnię. Pochłanianie promieni cieplnych odbywa się w powierzchniowych warstwach wody. Pozawidzialne długie promieniowanie (podczerwone o długości 7000Å i cieplne o fali dłuższej niż 8000Å) jest pochłaniane prawie w 99% w warstwie do głębokości 3-4 m. Jedynie cienka, powierzchniowa warstwa jest bezpośrednio nagrzewana promieniami słonecznymi. Przechodzenie ciepła w wodzie jest bardzo słabe - jest ona niezmiernie złym przewodnikiem ciepła. Proces nagrzewania wody z głębszych warstw zachodzi dzięki zjawisku konwekcji. Prądy konwekcyjne wytwarzane są na zasadzie zmian gęstości wody przy zmianach temperatury. Cząsteczki wody ogrzane do temperatury +4oC są cięższe od chłodniejszych i przemieszczając się w głąb stanowią o istocie pionowych prądów konwekcyjnych. Jednak zasadnicze znaczenie dla przemieszczania się cząsteczek wody i tym samym ogrzania głębszych warstw zbiorników wodnych, ma w wypadku wód stojących wiatr, a w przypadku rzek prąd wody.
Rozmieszczenia ciepła w zbiorniku jest specyficzne i uzależnione od elementów geograficznych, takich jak np.: szerokość geograficzna lub stopień wyniesienia nad poziom morza. Rozkład ciepła przybiera postać uwarstwienia termicznego. W strefie klimatu umiarkowanego jest ono wyraźnie wykształcone latem. Wraz ze zmianą warunków klimatycznych w ciągu roku następują 4 sezony limnologiczne (rys. 11).
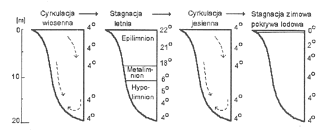
Rys. 11. Cykliczność roczna w termice wody jezior strefy umiarkowanej.
Wiosną, z chwilą zejścia pokrywy lodowej, woda mająca początkowo 0oC ogrzewa się pod wpływem promieniowania słonecznego. W wyniku prądów konwekcyjnych i mieszania wiatrowego przy temperaturze +4oC następuje wyrównanie gęstości wody w całym zbiorniku. Okres ten nazywa się przemieszaniem wiosennym lub cyrkulacją wiosenną, a zjawisko wyrównania termicznego - homotermią. W tym czasie następuje wyrównanie zawartości rozpuszczonych gazów i soli (elektrolitów) w całym słupie wody - dotąd gazy znajdowały się w warstwach powierzchniowych, a sole mineralne w przydennych. Zjawisko to ma kolosalne konsekwencje ekologiczne. Następuje wzbogacanie górnych warstw w sole biogenne, takie jak azotany i fosforany, dzięki którym może zachodzić produkcja biologiczna. Dalszy wzrost temperatury powoduje wytworzenie się pod powierzchnią zbiornika warstwy wody ciepłej, a więc o mniejszej gęstości (lżejszej), nie mającej tendencji do opadania, mieszanej tylko siłą wiatru. Warstwa ta stanowi oddzielną masę wodną w stosunku do leżącej pod nią wody zimnej o temperaturze niższej zwykle o kilkanaście stopni. Między tymi dwoma warstwami istnieje trzecia, o temperaturze pośredniej. Układ taki składający się z trzech warstw trwa przez całe lato, a nazywa się stagnacją letnią.
Górną warstwę wody ogrzanej, dobrze wymieszanej o podobnej temperaturze nazywa się epilimnionem. Leżącą poniżej cienką warstwę skoku termicznego, gdzie temperatura spada szybko wraz z głębokością (o jeden stopień na jeden metr), nazywa się metalimnionem lub termokliną,. Głębiej zalega woda, która latem nie podlega mieszaniu, a jej temperatura wynosi około +4oC. Warstwa ta, sięgająca do dna zbiornika, nazywa się hypolimnionem. Taki układ, tzn. gdy woda cieplejsza zalega pod powierzchnią jeziora, a zimna cięższa na dnie, nazywa się uwarstwieniem prostym. Gdy temperatura wody warstwy powierzchniowej nagrzeje się do ok. +10oC, wiatr nie jest już na ogół w stanie mieszać jej jako lżejszej z cięższą zimną wodą przydenną. Wówczas cyrkulacja wody w postaci prądów powierzchniowych i prądów powrotnych ścielących się ponad termokliną obejmuje tylko epilimnion.
Jesienią, gdy temperatura górnych warstw oziębi się i osiągnie wartość +4oC, nastąpi moment wyrównania termicznego w pionie. Jest to okres homotermii, czyli przemieszania jesiennego.
Zimą, gdy na powierzchni wody zalega lód, a temperatura pod nim wynosi 0oC, następuje okres stagnacji zimowej. Stosunki cieplne są wówczas odwrotne niż latem i nazywa się je uwarstwieniem odwróconym. Epilimnion jest zimą bardzo cienki, także cienka jest termoklina, która wykształca się płytko pod powierzchnią. Hypolimnion stanowi wówczas zasadniczą część mas wodnych. Przedstawiony schemat stosunków termicznych może w różnych sytuacjach wykazywać pewne odchylenie.
Wymieszanie wód zachodzi zwykle aż do dna. Mówimy wówczas o przemieszaniu zupełnym, czyli holomiksji. Jednak w warunkach szczególnych, np.: w jeziorach głębokich, osłoniętych od wiatru, dolne masy wody mogą nie podlegać krążeniu. Ma wówczas miejsce przemieszanie częściowe, czyli meromiksja. W tym przypadku górna warstwa wody podlegająca mieszaniu, zwana jest miksolimnionem, zaś ustabilizowana, stagnująca warstwa nosi nazwę monimolimnionu. Przyczyny meromiksji mogą być różne. W przypadku ograniczenia oddziaływania wiatru wskutek silnego osłonięcia zbiornika wodnego będzie to meromiksja mechaniczna. Gdy na dnie jeziora wypływają źródła o zimnej i bogatej w elektrolity wodzie i tym samym cięższej od warstw pozostałych, zachodzi meromiksja krenogeniczna. Na skutek działania organizmów dennych może zmieniać się skład chemiczny wody, np.: przy dnie nagromadzają się produkty procesów mikrobiologicznych, co w efekcie prowadzi do zwiększenia się gęstości wody; może więc zaistnieć meromiksja biogeniczna.
Siła oddziaływania wiatru i stopień osłonięcia zbiornika nie tylko wpływa na zasięg mieszania, ale również determinuje szybkość mieszania. Przy braku osłony i silnym oddziaływaniu wiatru zachodzi szybkie mieszanie się wody - tachymiksja. Natomiast przy bardzo słabym wietrze mieszanie jest niezmiernie powolne i nazywane jest bradymiksją. Zjawisko pośrednie do dwóch wymienionych to eumiksja.
Zróżnicowanie termiki jezior zależy przede wszystkim od położenia geograficznego i wyniesienia nad poziom morza. Ze względu na to kryterium Forel wyodrębnił 3 typy termiczne jezior:
- jeziora ciepłe o temperaturze wody stale wyższej lub co najmniej równej +4oC;
- jeziora strefy umiarkowanej o cyklu zmian sezonowych opisanych wyżej, z uwarstwieniem prostym w lecie i odwróconym w zimie;
- jeziora zimne o temperaturze przez cały rok niższej lub najwyższej równej +4oC.
Wskutek dalszych badań termiki jezior, szczególnie Afryki równikowej i Archipelagu Sundajskiego, uzyskano nowe dane pozwolające na wyodrębnienie grupy jezior tropikalnych, o wybitnej stabilności termicznej, tak w przekroju pionowym od powierzchni do dna, jak i w cyklu rocznym.
Przyjęty ostatnio w literaturze podział jezior wg. Hutchinsona i Löfflera (1956) wyróżnia sześć typów, zróżnicowanych pod względem częstotliwości mieszania w ciągu roku:
- Jeziora amiktyczne - stale zamarznięte, odcięte powłoką lodową od działania wiatru i promieniowania słonecznego. Nie wykazują zupełnie mieszania. Zalicza się do nich nieliczne jeziora polarne i wysokogórskie.
- Jeziora monomiktyczne zimne - o temperaturze nie przekraczającej +4oC. W zimie są pokryte lodem i mają uwarstwienie odwrócone, latem występuje cyrkulacja przy temperaturze +4oC. Są to zbiorniki typu polarnego, występujące także w wysokich górach, latem często mające kry lodowe na powierzchni.
- Jeziora dimiktyczne - o dwóch okresach przemieszania i uwarstwieniu prostym w lecie oraz odwróconym zimą. Są to jeziora strefy umiarkowanej oraz niższych partii wysokich gór strefy tropikalnej i subtropikalnej.
- Jeziora monomiktyczne ciepłe - o temperaturze wody nigdy nie spadającej poniżej +4oC. Występuje w nich jedno uwarstwienie o charakterze prostym i jedna cyrkulacja w sezonie chłodnym. Są to jeziora strefy subtropikalnej.
- Jeziora oligomiktyczne - o temperaturze stale powyżej +4oC. Woda jest ciepła na całym przekroju. Przemieszania są rzadkie i słabe, zwykle nieregularne. Są to jeziora wilgotnych rejonów tropikalnych, niezbyt wysoko wyniesione nad poziom morza.
- Jeziora polimiktyczne - o temperaturze niskiej, lecz zawsze powyżej +4oC, z licznymi w ciągu roku okresami przemieszania (nawet codziennymi). Są to zbiorniki płytkie, łatwo tracące ciepło, występujące zwykle w górzystych rejonach strefy tropikalnej.
Termika płytkich zbiorników wodnych wykazuje specyficzną charakterystykę. Nie mają one stałego i wyraźnego uwarstwienia. Wyróżnia się 3 typy takich zbiorników:
- Nieduże i płytkie zbiorniki o nietrwałym uwarstwieniu termicznym, takie jak: stawki, sadzawki i młaki. W dzień podczas silnego nasłonecznienia powstaje duża różnica między temperaturą na powierzchni a ponad dnem. Jednakże podczas chłodniejszej nocy wytwarza się homotermia i tym samym zachodzi całkowita cyrkulacja.
- Zbiorniki o niedużej powierzchni, ale stosunkowo głębokie, czyli charakteryzujące się dużą głębokością względną. Wykształca się tam latem normalne uwarstwienie proste, o podobnej różnicy temperatur między powierzchnią a dnem jak w głębokich jeziorach umiarkowanych.
- Zbiorniki płytkie, ale o bardzo dużej powierzchni, takie jak stawy lub rozległe jeziora strefy gorącej. Są to zbiorniki nagrzane do dna, o niedużych wahaniach dobowych.
Wody płynące charakteryzują się ruchem postępowym i obrotowym, co powoduje ciągłe ich mieszanie się. Tym samym wytworzenie się uwarstwienia termicznego jest utrudnione i wody te mają w zasadzie temperaturę równomierną od powierzchni do dna. Jednak w rzekach o bardzo powolnym prądzie mogą wytworzyć się w miejscach głębokich stosunki termiczne, z zaznaczonym uwarstwieniem termicznym, podobne do panujących w jeziorach. Najczęściej jednak różnica między temperaturą wody przy dnie a na powierzchni wynosi 1-2oC. Mimo takiej homotermii różnice w nagrzaniu wody w rzece są całkiem wyraźne. Latem woda jest najcieplejsza przy brzegach, najzimniejsza w środku nurtu; jesienią i zimą jest odwrotnie. Przyczyną tego jest ocieplający wpływ nagrzanych brzegów latem, a ochładzający zimą. Cechą charakterystyczną rzek jest nieduża roczna amplituda temperatur w ich górnym biegu, a znaczna w dolnym. Dotyczy to szczególnie tych rzek, które biorą początek ze źródeł górskich, z lodowców lub pól śnieżnych. W takim wypadku amplituda w odcinku wysokogórskim waha się od 1 do 6oC, w odcinku potokowym do 15oC, a w dolnym może dochodzić do 24oC. Temperatura wody źródeł jest prawie stała i podlega małym wahaniom. Tak więc woda źródlana wypływająca z głębszych warstw wodonośnych ma w zimie wodę ciepłą, a w lecie zimną.
Termika mórz, a szczególnie oceanów jest mniej zbadana niż wód śródlądowych. Prosty układ stosunków termicznych podobny do jezior typu umiarkowanego panuje w morzach wewnętrznych, tak jak jest to np. w Bałtyku. Występuje w nim letnia i zimowa stagnacja oraz wiosenna i jesienna cyrkulacja. Ze względu na zaleganie głębiej wód bardziej zasolonych, czyli cięższych, zasięg cyrkulacji termicznej jest ograniczony do górnych warstw. Bałtyk ma więc specyfikę zbliżoną do jezior meromiktycznych (rys. 12). Inaczej przedstawiają się stosunki termiczne w głębokim i równomiernie zasolonym Morzu Śródziemnym, położonym w strefie klimatu ciepłego. Tam temperatura wody na głębokości 150-200 m osiąga około 14oC. Głębiej niewiele spada osiągając ponad dnem 12,5-13,5oC, co odpowiada mniej więcej temperaturze na powierzchni zimą. Latem woda ogrzewa się na powierzchni do ponad 25oC.
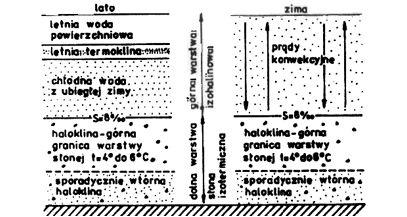
Rys. 12. Uwarstwienie termohalinowe w Bałtyku
W oceanach, w związku z różnym stopniem nagrzania powierzchni wody zależnym od kąta padania promieni słonecznych, roczna amplituda temperatur powierzchniowych jest różna. Na biegunach i na równiku nie przekracza kilku stopni, natomiast w pasie pośrednim dochodzi do 10oC. Wahania te ograniczone są do warstwy wód powierzchniowych sięgającej do około 200 m głębokości. Głębiej zalegają wody, których temperatura stopniowo spada, osiągając w głębinach prawie 0oC. Jest to związane ze specyfiką krążenia mas wodnych w oceanach i istnieniem prądów głębinowych, niosących zimne wody polarne ponad dnem oceanicznym. Zaproponowany przez Defanta schemat stosunków termicznych Oceanu Atlantyckiego zakłada pewną analogię do jeziornego uwarstwienia latem, gdyż ciepłe wody strefy zwrotnikowej odpowiadać mogą epilimnionowi, zaś zimne, polarne i głębinowe hypolimnionowi. Rozdzielone są one tzw. frontem polarnym, czyli warstwą stykową, która zapobiega mieszaniu się w pionie wód o różnej temperaturze i tym samym różnej gęstości. Schemat krążenia wód wg Defanta (rys. 13) zakłada cyrkulację zamkniętą oddzielnie w ramach obu półkul. Jest to duże uproszczenie, wiadomo bowiem, iż cyrkulacja w Oceanie Atlantyckim jest bardziej skomplikowana. Jednak przyjęcie tego modelu pozwoliło na wyjaśnienie faktu znajdowania się przy dnie oceanów wody o temperaturze około 0oC, a nie +4oC, czego należałoby się spodziewać po uwzględnieniu prawidłowości termicznych wód naturalnych.
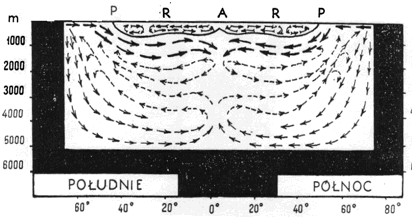
Rys. 13. Schemat krążenia wód oceanicznych w Atlantyku wg Defanta. A - równik, P - front polarny (granica między wodą ciepłą a zimną), R - rejon prądów ciepłych
Na powierzchni wszechoceanu można więc wyróżnić pas wód gorących oraz dwa rejony zimne, polarne, a także dwie strefy pośrednie, czyli pięć pasów klimatycznych. Znaczne zróżnicowanie tych pasów, a co za tym idzie zróżnicowanie warunków życia, było podstawą wyodrębnienia pięciu krain biogeograficznych: jednej ciepłowodnej - tropikalnej, dwóch zimnowodnych - arktycznej i antarktycznej, oraz dwóch umiarkowanych - borealnej na północy i notalnej na południu (rys. 14). Granica między nimi nie przebiega ściśle równoleżnikowo. Związane jest to z nierównoległym przebiegiem prądów powierzchniowych. Tak więc najlepszym wyznacznikiem zasięgu danej krainy są tzw. formy przewodnie, a więc organizmy o wąskim przystosowaniu termicznym.
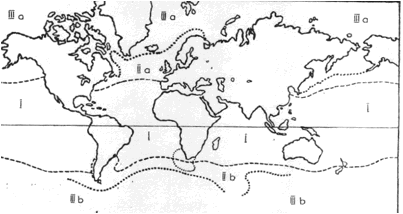
Rys. 14. Krainy biogeograficzne w morzach: I - ciepłowodna; II - umiarkowane: a - borealna, b - notalna; III - zimnowodne: a - arktyczna, b - antarktyczna
Formami przewodnimi w strefie ciepłowodnej są ciepłolubne mięczaki skrzydłonogie Pteropoda, prócz tego ryby latające, a przede wszystkim rafy koralowe i formacje namorzyn, zaś z planktonu wiciowce wapienne, krzemionkowe i bruzdnice. W morzach polarnych zaznacza się przewaga okrzemek, a najbardziej charakterystycznym składnikiem fauny polarnej są obunogi Amphipoda i równonogi Isopoda. Dla krainy wód arktycznych typowymi są również przedstawiciele skorupiaków - widłonogi Calanus hyperboreus i lasonogi Mysidacea. Wody antarktyczne charakteryzuje obecność kikutnic i kryla (skorupiaki z grupy Euphausiacea), ryby białokrwiste reprezentowane przez Notothenidae oraz pingwiny. Morza umiarkowane stanowią środowisko przejściowe i dlatego w ich faunie jest obecnych sporo grup typowych zarówno dla rejonów biegunowych, jak i krainy tropikalnej. Za formę przewodnią dla krainy borealnej w Atlantyku można uznać kulistą zielenicę planktonową Halosphaera viridis.
Chemizm wód
Woda w przyrodzie nie występuje nigdy jako czysty związek chemiczny, a to dlatego, że jest ona doskonałym rozpuszczalnikiem gazów, cieczy i ciał stałych. Wynika to ze specyfiki jej budowy molekularnej, o czy była mowa w poprzednim rozdziale.
Stopień rozpuszczalności substancji chemicznych zależy od stałej rozpuszczalności i od temperatury. Dla ciał stałych zależność od temperatury jest wprost proporcjonalna, natomiast dla gazów odwrotnie proporcjonalna, tzn. w wyższej temperaturze rozpuszczalność gazów maleje.
Stosunki chemiczne w wodzie naturalnej są ściśle powiązane z zachodzącymi w niej procesami biologicznymi. Dotyczy to szczególnie warstwy powierzchniowej, gdzie rośliny pobierają z wody sole mineralne, wytwarzają z nich substancje organiczne będące budulcem komórek i tkanek. Równocześnie rośliny i zwierzęta wydzielają do wody produkty przemiany materii, które zwykle są prostymi związkami chemicznymi. Obumarłe organizmy, wskutek działalności bakterii, zostają rozkładane do prostych soli mineralnych, które znowu wracają do obiegu.
Gazy w środowisku wodnym
Gazy w wodzie naturalnej pochodzą z trzech źródeł: z atmosfery, są produktem procesów życiowych organizmów albo wytworem przemian chemicznych zachodzących w samej wodzie.
Z atmosfery przedostaje się do wody tlen, azot, dwutlenek węgla i gazy szlachetne. W samej wodzie na skutek procesów biologicznych i chemicznych wytwarza się, poza trzema wyżej wymienionymi, siarkowodór, wodór, amoniak, dwutlenek siarki oraz tlenek węgla. Zawartość w wodzie trzech podstawowych gazów - azotu, tlenu i dwutlenku węgla - jest dużo mniejsza niż w atmosferze (tab. 1). Zależy ona od temperatury i zasolenia, choć procentowy ich udział jest prawie stały, co z punktu widzenia zjawisk biologicznych jest niezmiernie ważne.
Tabela 1. Zawartość trzech podstawowych gazów w różnych środowiskach
Środowisko N2 O2 CO2 Razem
Atmosfera cm3/l 790 210 0,3 1000
% 79 21 0,03 -
Woda słodka 0oC cm3/l 18,64 10,29 0,51 29,4
% 63 35 1,7 -
20oC cm3/l 12,59 6,57 0,26 19,4
% 65 34 1,3 -
Woda morska 0oC cm3/l 11,42 8,04 0,44 19,9
% 63 35 1,9 -
20oC cm3/l 10,42 5,36 0,23 16,0
% 65 34 1,4 -
Źródło: Lityński A. Hydrobiologia ogólna. Warszawa, PWN 1952.
Tlen
Rozpuszczony w wodzie naturalnej tlen pochodzi z dwóch źródeł: z atmosfery (drogą dyfuzji) oraz z procesu biologicznego, jakim jest fotosynteza.
Przenikanie tlenu z atmosfery przez gładką powierzchnię zbiornika wodnego jest procesem mało efektywnym. Stwierdzono, że w wodzie pozbawionej tlenu, w warunkach stałej temperatury znalazłby się on na głębokości 6 m dopiero po roku i to zaledwie w stężeniu 0,25 cm3/l. W rzeczywistości proces przenikania tlenu do wody jest znacznie szybszy dzięki falowaniu. Na rozprzestrzenianie się tlenu w wodzie wpływa turbulencja i prądy.
Nasycenie wody naturalnej tlenem uzależnione jest od temperatury. Uwarstwienie termiczne powoduje, że jest ono różne na różnych głębokościach. Natomiast w okresach przemieszania jest na ogół w całym przekroju takie samo. Teoretyczne nasycenie tlenem wody w temperaturze 4oC wynosi około 13 mg, czyli 9 cm3/l. W okresie stagnacji zimowej rozpuszczonego tlenu jest więcej pod powierzchnią lodu (temp. poniżej 4oC) niż w warstwach głębszych. Odwrotnie powinno być podczas stagnacji letniej. W epilimnionie, gdzie panują wysokie temperatury, tlenu powinno być najmniej. Jest jednak inaczej, a to dlatego, że w warstwie tej zachodzi proces fotosyntezy stanowiący podstawowe źródło tlenu. Ilość tlenu wyprodukowanego w tym procesie zależy od zagęszczenia roślin i natężenia światła. Produkcja tlenu w zbiorniku zachodzi w jego prześwietlonych strefach, a więc w płytkiej strefie przybrzeżnej (litoral), oraz w strefie wody otwartej (pelagial). W pierwszej z nich tlen produkują rośliny naczyniowe, glony bentosowe oraz fitoplankton, natomiast w drugiej wyłącznie fitoplankton.
Natężenie procesu fotosyntezy wykazuje wyraźną cykliczność dobową i sezonową.
W ciągu dnia jest on najintensywniejszy w godzinach popołudniowych. Nocą proces ten jest prawie zupełnie zahamowany. Fotosynteza jest intensywniejsza latem niż w innych sezonach. Podobnie kształtuje się nasycenie wody tlenem. W rejonach dużego zagęszczenia roślin, czy to w litoralu, czy to w toni wodnej, może zaistnieć przesycenie wody tlenem. Stan taki jednak szybko zanika - nadwyżka tlenu uchodzi do atmosfery.
Stan, w którym w wodzie, w danych warunkach fizycznych (szczególny wpływ ma temperatura), zawarta jest największa możliwa ilość tlenu nazywa się stanem nasycenia. Ilość tlenu potrzebna do uzyskania stanu nasycenia jest różna w różnych warunkach środowiskowych, a do jej wyrażenia stosuje się zawartość bezwzględną gazu lub procent nasycenia. Procent nasycenia jest to stosunek rzeczywistej zawartości do stanu nasycenia, np. woda zawiera 50% tlenu wtedy, gdy zawartość O2 jest tam o 50% mniejsza niż zawartość możliwa w stanie nasycenia przy tej samej temperaturze i ciśnieniu oraz takim samym zasoleniu. Różnicę między możliwym stanem nasycenia a rzeczywistą zawartością tlenu określa się jako deficyt tlenowy. Jest on również, tak jak zawartość tlenu, wyrażony w procentach lub w cm3 albo w mg na litr.
Deficyt tlenowy, czyli ubytki tlenu, jest spowodowany kilkoma procesami zachodzącymi w środowisku wodnym. Spośród nich podstawowym procesem, podczas którego odbywa się konsumpcja tlenu, jest oddychanie. Proces ten trwa dniem i nocą. Dniem jest on rekompensowany przez fotosyntezę. Drugi proces, powodujący spadek zawartości tlenu w wodzie, to rozkład substancji organicznej, inaczej utlenianie. Jest to proces zależny od temperatury. Stąd jego tempo będzie szybsze latem w epilimnionie, a wolniejsze w głębi zbiornika. Tlen bierze udział w wielu reakcjach chemicznych, w efekcie których może być związany na stałe w pewnych solach, np. rozpuszczalne w wodzie sole żelazawe wiążą tlen przechodząc w słabo rozpuszczalne sole żelazowe. Ponadto tlen może mechanicznie uchodzić do atmosfery w wyniku tzw. banieczkowania, spowodowanego chociażby gwałtownymi zmianami temperatury lub unoszeniem się do atmosfery w postaci banieczek tlenu zmieszanego z innymi bardziej rozpuszczalnymi gazami.
Zawartość tlenu w wodzie jest więc efektem bilansu procesów produkcyjnych i konsumpcyjnych. Można ją, podobnie jak rozkład temperatury, przedstawić graficznie (rys. 15). Przebieg krzywej zawartości tlenu zależy od żyzności zbiornika, a tym samym intensywności produkcji i rozkładu materii organicznej na danej głębokości.
W zbiornikach żyznych różnice w zawartości tlenu między powierzchnią a dnem mogą być duże. Zawartość tlenu, podobnie jak temperatura, jest ważnym czynnikiem ekologicznym. Każdy gatunek ma swoje optimum oraz zakres warunków tlenowych, które może znieść. Organizmy o wysokich wymaganiach tlenowych często przystosowane są do życia w niskich temperaturach, zaś znoszące duże wahania temperatury, czyli eurytermiczne, wytrzymują na ogół duże niedobory tlenowe.
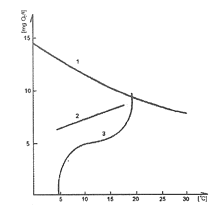
Rys. 15. Zawartość tlenu 1 - w wodzie destylowanej (rozpuszczalność fizyczna) 2 - w wodzie naturalnej w jeziorze ubogim 3 - w jeziorze żyznym
Zróżnicowanie rozkładu pionowego zawartości tlenu w wielu zbiornikach było powodem, dla którego Thienemann wyodrębnił dwa główne typy tlenowe jezior: podalpejski i bałtycki (rys. 16). Różnice dotyczą zawartości tlenu w hypolimnionie.
Typ podalpejski charakteryzuje duża zawartość tlenu we wszystkich warstwach, zaś w jeziorach typu bałtyckiego w hypolimnionie, a zwykle i w metalimnionie, zaznacza się deficyt tlenowy, który przy dnie może oznaczać całkowity brak tlenu. Uwarstwienie tlenowe nawiązuje do uwarstwienia termicznego, a także warstwowego zasięgu światła.
Tak więc te trzy czynniki ekologiczne w okresie stagnacji letniej są optymalne w warstwie powierzchniowej, gdzie zachodzi produkcja. Warstwa ta nazywana jest trofogeniczną, czyli wytwórczą. Głębiej natężenie procesów produkcyjnych spada, a większą rolę zaczynają odgrywać procesy rozkładu. Głębokość, na której te dwa typy procesów się równoważą (produkcja tlenu i jego ubytki są zbilansowane) nazywa się poziomem kompensacyjnym. W przybliżeniu określany jest on poprzez odczyt widzialności krążka Secchiego pomnożony przez 1,5. Poniżej rozpościera się warstwa wody, gdzie rola producentów jest ograniczona i procesy zużycia tlenu przewyższają procesy jego produkcji. Jest więc to warstwa rozkładu materii organicznej zwana inaczej warstwą trofolityczną.
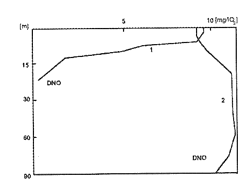
Rys. 16. Rozkład pionowy zawartości tlenu 1 - w jeziorze żyznym (typ bałtycki) 2 - w jeziorze ubogim (typ podalpejski)
Uwarstwienie tlenowe notowane jest również w morzach i oceanach. W Atlantyku w strefie międzyzwrotnikowej górna warstwa prześwietlona zawiera znaczne ilości tlenu, głębiej zawartość bardzo znacznie spada, nieraz nawet poniżej 1 cm3/l. Natomiast wody poniżej 1500 m zawierają około 5-6 cm3/l. Zupełnie inaczej kształtuje się uwarstwienie tlenowe w wodach arktycznych. W warstwach górnych jest prawie 100% nasycenia i zauważalny jest równomierny spadek w kierunku dna, zachodzący bardzo wolno, gdyż nawet na głębokości ponad 3000 m deficyt wynosi zaledwie 15%. Różne rejony wszechoceanu mają swoją specyfikę uwarstwienia tlenowego wynikającą z różnorodności warunków środowiskowych. Są rejony, a dotyczy to szczególnie mórz izolowanych, gdzie deficyt tlenowy w warstwach głębszych jest bardzo duży i jak np. w Morzu Czarnym, może osiągnąć 100%. Warstwa dobrze natleniona w tym morzu sięga do około 150 m, głębiej ilości tlenu są znikome lub go brak. W jego miejsce pojawia się w dużych ilościach siarkowodór. Takie uwarstwienie tlenowo-siarkowodorowe jest wynikiem braku cyrkulacji pionowej, spowodowanej zaleganiem poniżej granicy występowania tlenu mas wodnych o wyraźnie wyższym zasoleniu. Natomiast w przylegającym do Morza Czarnego płytkim Morzu Azowskim, gdzie również okresowo ponad dnem tworzy się siarkowodór, prawie każdy większy wiatr powoduje wymieszanie wód do dna. W ten sposób okresy uwarstwienia tlenowego tworzące się w okresach ciszy są krótkotrwałe i są zastępowane okresami homotermii, a tym samym homooksygenii, czyli wyrównania różnic w zawartości tlenu w pionie. Także w Bałtyku od wielu lat stwierdza się duże obszary dna z zalegającym siarkowodorem. Są to tzw. pustynie azoiczne - tam gdzie występuje siarkowodór brak jest tlenu, stąd formy życia tlenowego w tych rejonach są niemożliwe. Powiększanie się tych obszarów jest ograniczane przez specyficzne dla Bałtyku zjawisko wlewów przez Cieśniny Duńskie dobrze natlenionej wody z Morza Północnego. Woda ta jako bardziej słona od wody bałtyckiej ściele się po dnie, odświeżając warstwy przydenne.
Dwutlenek węgla
Drugim co do ważności biologicznej gazem rozpuszczonym w wodzie jest dwutlenek węgla. Pochodzi on głównie z procesów fermentacyjnych zachodzących w mule dna, z oddychania zwierząt i w niedużej ilości z rozpuszczania się w wodzie gazowego CO2 zawartego w powietrzu. W atmosferze jest go niewiele, zaledwie 0,03-0,04%, co powoduje, że jego ciśnienie parcjalne jest niewielkie. W wodzie CO2 jest znacznie więcej: około 0,6%. Ilość CO2 w wodzie uzależniona jest od intensywności fotosyntezy i niekiedy w pogodne, ciepłe dni już w godzinach przedpołudniowych bywa wyczerpany cały zapas wolnego dwutlenku węgla. Wahania dobowe CO2 w wodzie mogą być bardzo duże. Są one tym większe, im mniejsza jest zawartość wapnia w wodzie. Związane są z tym również wahania odczynu wody (pH) - fluktuacje zawartości dwutlenku węgla i wapnia oraz wartości pH są ze sobą ściśle powiązane.
Dwutlenek węgla rozpuszczony w wodzie wchodzi w połączenia z wodą dając słaby kwas węglowy H2CO3, który szybko ulega dysocjacji na jony H+ i HCO3- . Jon HCO3- łatwo reaguje z jonami metali lekkich, szczególnie z wapniem, dając dwuwęglan wapnia Ca(HCO3)2 rozpuszczający się wodzie w ilości 1170 mg/l. Jest to jednak związek nietrwały, łatwo rozpadający się z wytworzeniem gorzej rozpuszczającego się w wodzie węglanu wapnia CaCO3, którego rozpuszczalność w wodzie wynosi jedynie 13 mg/l:
Ca(HCO3)2 <=> CaCO3 + CO2 + H2O
Utrzymanie się dwuwęglanu wapnia w roztworze uzależnione jest od ilości CO2. Gdy jest ona odpowiednia wówczas ustala się pewien stan równowagi. Dwutlenek węgla powodujący tę równowagę, czyli potrzebny do utrzymania dwuwęglanu wapnia w roztworze, nazywany jest CO2 równowagi lub CO2 przynależny.
W wodach naturalnych w związku z procesami biologicznymi występują wahania zawartości dwuwęglanu wapnia. Wahania te są ściśle związane z ilością dwutlenku węgla. Rozkład dwuwęglanu odbywa się głównie w trakcie procesu fotosyntezy roślin, które w dzień wykorzystują go w dużych ilościach. Powodują one przez to zmniejszenie się ilości CO2 przynależnego, a w efekcie rozpad dwuwęglanu wapnia z wytrąceniem węglanu wapnia, mogącego istnieć w roztworze tylko w ilości do 13 mg/l. Tak więc cały nadmiar wytrąca się z wody i osadza w postaci drobnego pyłu najczęściej na powierzchni roślin. Wytrącanie się stałego węglanu wapnia nazywa się biologicznym odwapnianiem wody, gdyż jest efektem biologicznej działalności roślin wodnych. W trakcie tego procesu wytrąca się również częściowo kwas fosforowy rozpuszczony w wodzie, który wchodzi w reakcje z wytwarzającym się węglanem wapnia, dając trudno rozpuszczalny fosforan wapniowy opadający na dno.
3CaCO3 + 2KH2PO4 => Ca3(PO4)2 + 2KHCO3 + CO2 + H2O
Podczas tej reakcji dwutlenek węgla zostaje uwolniony i może więc być dalej biologicznie użyteczny. Równocześnie jednak następuje zubożenie wody w fosfor, będącym z kolei elementem biologicznym niezbędnym dla procesów życiowych.
Innym procesem biologicznym, który wpływa na wahania zawartości CO2 w wodzie jest oddychanie. Szczególnie nocą, gdy ustaje fotosynteza, oddychanie może doprowadzić do znacznego zwiększenia ilości CO2. Gdy ilość dwutlenku węgla jest większa niż CO2 przynależnego to nadwyżka taka powoduje rozpuszczanie węglanu wapnia i przenoszenie go do roztworu w formę dwuwęglanu. Jest więc to jakby ponadprzynależna ilość CO2. Nazywa się go często CO2 agresywnym ze względu na rozpuszczanie kamieni wapiennych na dnie lub brzegach, a także korozję umocnień betonowych.
Dwuwęglan wapnia jako sól słabego kwasu jest bardzo ważnym regulatorem (buforem) odczynu wody, zapobiegającym silniejszym wahaniom pH. W obecności mocnych kwasów lub zasad wchodzi z nimi w reakcję, zobojętniając je.
Ca(HCO3)2 + 2HCl => CaCl2 + 2CO2 + 2H2O
Ca(HCO3)2 + 2KOH => CaCO3 + K2CO3 + 2H2O
Podczas tych reakcji nie następuje uwolnienie ani jonów H+, ani OH-, od których proporcji zależy odczyn wody. Tak więc pośrednio na odczyn wody wpływa dwutlenek węgla. Z odpowiednimi wartościami pH wody wiąże się obecność danej postaci CO2 w wodzie (rys. 17).
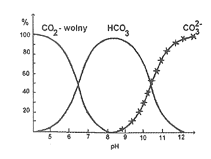
Rys. 17. Procentowy udział różnych form dwutlenku węgla w zależności od pH wody
Przy pH około 4 dwutlenek węgla występuje w 100% w postaci wolnej, która znika prawie zupełnie już przy pH=8. Inaczej zachowuje się jon HCO3-, który wraz ze wzrostem pH wymienia się z wolnym CO2, aby go w pełni zastąpić przy pH=8. Natomiast przy dalszym wzroście pH jego zwartość maleje, a jego miejsce zajmuje jon CO3--.
Jak już wspomniano wyżej, przed zbyt silnymi wahaniami odczynu zabezpiecza wodę wapń. Jest więc on bardzo ważnym czynnikiem w procesie produkcji biologicznej w wodzie. Wody o małej zawartości wapnia są zwykle zakwaszone i nieurodzajne. Wody o zawartości wapnia powyżej 65 mg/l CaCO3 są silnie buforowane o wahaniach odczynu w granicach 2,5 stopnia pH. Duże zapasy CO2 związanego w węglanach zapewniają dużą produkcję roślinną, tym samym dużą urodzajność. Odczyn wody jest czynnikiem ekologicznym o istotnym znaczeniu w biologii poszczególnych gatunków. Organizmy wodne nie znoszą dużych wahań odczynu wody. Dla większości optymalny zakres pH mieści się pomiędzy 6,5 a 8,5. Woda w warstwie powierzchniowej oceanów i wielu otwartych mórz ma odczyn zasadowy od pH 7,8 do 8,4 lub nawet 8,0-8,3. Większe jeziora o normalnej zawartości węglanów mają także wodę zasadową o pH 8 lub nieco wyższym. Podobnie kształtuje się odczyn wody w wielu większych rzekach. Jednakże istnieją również zbiorniki wodne, w których poprzez nagromadzenie wolnego CO2 lub znaczną zawartość kwasów mineralnych lub organicznych (np. humusowych) odczyn wody jest mniej lub bardziej kwaśny.
Skoro więc zawartość dwutlenku węgla (podobnie jak tlenu) uzależniona jest od procesów biologicznych zachodzących w zbiorniku wodnym to wykazywać musi ona podobną wyraźną cykliczność dobową (rys. 18) i zmienność sezonową. Ta ostatnia odnosi się do rozkładu pionowego. W normalnie żyznych jeziorach w okresie stagnacji letniej dwutlenek węgla zachowuje się odwrotnie niż tlen. Jego stężenie wzrasta wraz z głębokością i spadkiem temperatury (rys. 19).
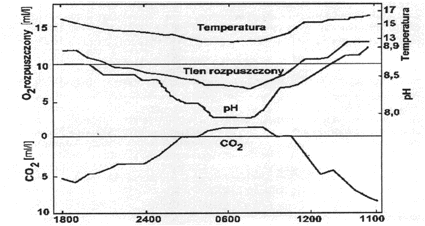
Rys. 18. Zmienność dobowa zawartości dwutlenku węgla oraz innych parametrów w wodzie powierzchniowej latem.
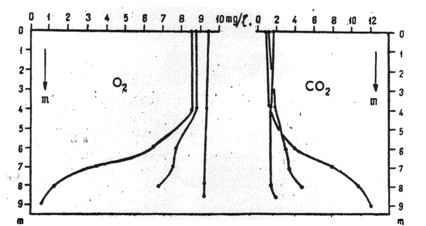
Rys. 19. Rozkład pionowy stężenia dwutlenku węgla i tlenu na przełomie przemieszania wiosennego i uwarstwienia letniego.
Sole mineralne
W wodach naturalnych mogą być obecne teoretycznie wszystkie pierwiastki i związki chemiczne notowane w skorupie ziemskiej. Jest tak w przypadku wody morskiej. Jednak skład chemiczny wód śródlądowych, z nielicznymi wyjątkami, jest odmienny i charakteryzuje się znacznie mniejszym stopniem stężenia. Ponadto występowanie dominujących jonów nadaje tym wodom inny charakter niż wodom morskim (rys. 20). W wodzie morskiej w największej ilości występuje sól kuchenna, podczas gdy wody słodkie charakteryzuje przewaga węglanów nad innymi solami.
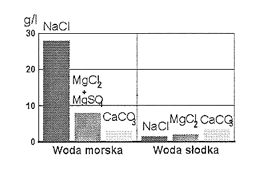
Rys. 20. Zawartość podstawowych soli w 1 litrze wody morskiej i słodkiej.
Zasolenie rozumie się jako całkowitą ilość rozpuszczonych elektrolitów w litrze wody. Stopień zasolenia podaje się w promilach i oznacza symbolem S o/oo. Zasolenie S=10o/oo oznacza, że w 1 litrze wody rozpuszczonych jest 10 g soli. Stopień zasolenia wód jest zróżnicowany. Można go uszeregować od zbiorników ze śladową ilością soli do zbiorników o bardzo dużym jej stężeniu. Różne są w literaturze naukowej systemy klasyfikacji wód ze względu na ich zasolenie. Powszechnie przyjęty jest system wenecki, który wyróżnia cztery zasadnicze kategorie wód (tab. 2).
Tabela 2.
Kategorie wód w zależności od stopnia zasolenia (wg systemu weneckiego).
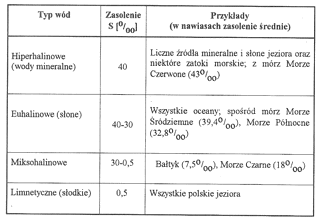
Wody miksohalinowe (słonawe) zostały dalej podzielone na trzy podkategorie: wody miksopolihalinowe, o zasoleniu 30-18o/oo, miksomezohalinowe - 18-5o/oo i miksooligohalinowe - 5-0,5o/oo.
Bałtyk jest typowym morzem słonawym o zmniejszającym się zasoleniu w kierunku północno-zachodnim. W rejonie Cieśnin Duńskich zasolenie wód powierzchniowych wynosi 19-20o/oo, w okolicach wyspy Rugia 8,5o/oo, przy Helu 7,75o/oo, w Zatoce Botnickiej 2-5o/oo. Tak duża gradacja zasolenia ma swoje konsekwencje biologiczne. Jedną z nich jest ubożenie gatunków flory i fauny morskiej w miarę obniżania się stopnia zasolenia (tab. 3). Inną jest karlenie form morskich związane ze spadkiem zasolenia.
Tabela 3.
Liczba gatunków flory i fauny bałtyckiej w zależności od stopnia zasolenia
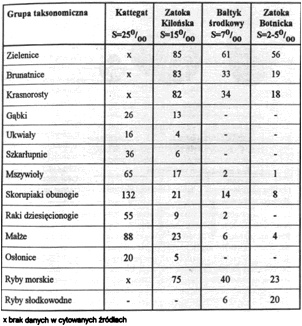
Źródło: Demel K. Życie morza. Wyd.4. Gdańsk, Wyd.Morskie 1974.
Pankow H., Algenflora der Ostsee. 1. Benthos. Jena, Fisher Verlag 1971.
Zjawisko karlenia widać szczególnie wyraźnie w przypadku małży (rys. 21). U mięczaków morskich żyjących w obniżonym zasoleniu, poza zmniejszoną wielkością ciała, zaznacza się również zmniejszenie grubości muszli. Wskazywałoby to na fakt, iż niższe zasolenie obniża tempo metabolizmu.
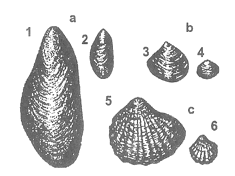
Rys. 21. Porównanie wielkości małży z rejonów o różnym zasoleniu a - omułek jadalny (Mytilus edulis): 1 - z Morza Północnego, 2 - z przybrzeżnych wód Finlandii; b - rogowiec bałtycki (Tellina baltica): 3 - z Zatoki Kilońskiej, 4 - z południowych przybrzeżnych wód Bałtyku; c - sercówka (Cardium edule): 5 - z Morza Północnego, 6 - z przybrzeżnych wód Finlandii
Zawartość elektrolitów jest więc różna w zależności od typu zbiornika. Może się także zmieniać w czasie. Elektrolity dostają się do wód w wyniku procesu rozpuszczania podłoża zbiornika. Wnoszone są także z dopływami i z opadami atmosferycznymi. Odzyskiwane są także w procesie rozkładu obumarłych organizmów. Zawartość elektrolitów w wodzie określa się ważąc pozostałość po spaleniu przefiltrowanej i odparowanej próby uzyskanej z określonej objętości wody. Można również pośrednio określić ogólną zawartość elektrolitów mierząc przewodnictwo elektrolityczne wody.
Zawartość soli mineralnych w wodach słodkich nie jest duża i waha się w granicach 50-600 mg/l. Najuboższe są zbiorniki wykształcone na podłożu skalistym, np. granitowym (ok. 50 mg/l), bogatsze są jeziora górskie, ale o podłożu wapiennym (100-200 mg/l). Jeszcze więcej soli jest w jeziorach niżowych (200-400 mg/l). W rzekach zawartość soli jest podobna (50-600 mg/l). Zbiorniki wykształcone na podłożu torfowym, tzw. wody kwaśne, zawierają bardzo mało elektrolitów, za to dużo materii organicznej. Zawartość elektrolitów w wodzie decyduje o jej jakości, co określa się mianem twardości ogólnej. Woda miękka zawiera poniżej 50 mg soli w litrze, większe wartości soli odnoszą się do wód średnio twardych i twardych.
Prawie wszystkie pierwiastki występujące jako sole rozpuszczalne w wodach naturalnych mają znaczenie biologiczne. Jedne z nich np. węgiel, azot, fosfor, siarka, magnez wapń, potas, żelazo, krzem, zużywane są przez rośliny w dużych ilościach. Inne zaś potrzebne są w bardzo małych ilościach. Są to tzw. mikroelementy, jak np. bar, mangan, molibden, miedź, brom, jod, fluor, glin, cynk. Obecność pierwszych działa stymulująco na wzrost roślin, drugie zaś w większych ilościach go hamują. Specyficzną rolę w środowisku odgrywają związki tlenowe azotu i fosforu, jako elementy biogenne (biogeny, nutrienty).
Azot i fosfor
Azot i fosfor są obok węgla najważniejszymi elementami budulcowymi żywej materii. Rośliny zużywają w dużej ilości sole kwasu azotowego i sole amonowe. Bardzo rzadko natomiast korzystają z wolnego azotu rozpuszczonego w wodzie. Wolny azot bywa pobierany przez niektóre bakterie, sinice oraz glony, przez co zwiększa się ogólna pula tego pierwiastka w wodzie. Azot w wodzie pochodzi niemal wyłącznie z atmosfery. Źródłem wolnego azotu w wodzie mogą być również bakteryjne procesy redukcji amoniaku. Z ogólnej zawartości azotu w wodzie prawie 50% przypada na związki organiczne, w tym do 80% mogą to być związki typu białkowego, takie jak proteiny, polipeptydy i aminokwasy. Wchodzą one w skład żywych organizmów i mogą być w stanie rozpuszczonym w wodzie jako efekt procesów rozkładu. Cykl przemian azotowych w wodzie jest rozbudowany przez obecność w nim różnego typu bakterii (rys. 22).
Rys. 22. Schemat cyklu krążenia azotu w wodzie
Ogólna zawartość azotu w wodach naturalnych jest zróżnicowana. Zależy to od typu zbiornika, rejonu zbiornika i głębokości. Zmienia się także w związku z porą roku. W wodach powierzchniowych oceanu za średnią przyjmuje się ok. 10 mg/m3 azotu. W rejonach otwartych azotu jest zazwyczaj mniej, a w basenach przybrzeżnych więcej (np. w Morzu Północnym powyżej 15 mg/m3). W rozmieszczeniu pionowym zaznacza się wyraźny wzrost wraz z głębokością. W Atlantyku u wybrzeży francusko-hiszpańskich w warstwie 0-50 m stwierdzono 90 mg N/NO3 w 1 m3, na głębokości 200 m - 150, 400 m - 230, a na 2000 m aż 325 mg. Wyjątkowo duże ilości azotu stwierdzono w głębszych warstwach Morza Czarnego, gdzie ogólna jego zawartość, głównie w postaci amoniaku, wynosi 1500 mg/m3. Jest to oczywiście związane z brakiem cyrkulacji pionowej i brakiem możliwości jego biologicznego zużytkowania. Podobne stosunki panują w głębokich jeziorach. I tak np. w jeziorze Górnym Lunz na powierzchni stwierdzono 1 mg N/NO3 w 1 m3, natomiast nad dnem 820 mg. Ilość w jeziorach górskich azotu jest bardzo mała. Znacznie więcej jest go w jeziorach niżowych, np. w hypolimnionie latem może być go ponad 1000 mg w 1 m3 (azot amoniakalny). W rzekach zawartość azotu jest różna w zależności od odcinka biegu rzeki - w dolnym jego ilości mogą być znaczne.
Zmienność sezonowa zawartości azotu w wodzie jest ściśle związana z cyklicznością sezonową w rozwoju fitoplanktonu. W strefie umiarkowanej, niezależnie czy w morzach, czy w zbiornikach śródlądowych, następuje ostry spadek stężenia azotu w wodzie tuż po zakwicie wiosennym. W okresie letnim wegetacja jest na ogół nieco mniejsza, co powoduje nieznaczne odnowienie zapasów naruszanych niekiedy przez jesienny, znacznie zresztą mniejszy od wiosennego, zakwit okrzemek. W okresie zimy, kiedy wegetacja jest zahamowana, następuje odbudowanie zapasów rozpuszczonych soli azotowych, które znowu zostaną wykorzystane na wiosnę.
Fosfor, podobnie jak azot, występuje w wodzie w związkach o różnej postaci: jako rozpuszczalny fosfor fosforanowy, rozpuszczalny fosfor organiczny i fosfor organiczny związany. Fosfor jest pierwiastkiem regulującym produkcję biologiczną i jako przenośnik energii jest elementem nieodzownym w procesach życiowych. Pierwiastek ten występuje w małych ilościach w wodzie naturalnej, a jego niedobór może ograniczać lub hamować produkcję. W porównaniu do azotanów, fosforany występują w wodach w znacznie mniejszych ilościach (0,001-0,1 mg/l P2O5). Są jednak także w mniejszym stopniu zużywane przez rośliny.
Rozmieszczenie fosforanów w zbiorniku wodnym jest na ogół odwrotnie proporcjonalne do stężenia tlenu. W epilimnionie fosfor jest szybko zużywany przez fitoplankton, natomiast wobec braku tlenu w hypolimnionie, sole żelazowe kwasu fosforowego ulegają redukcji na łatwiej rozpuszczalne żelazawe. Dzieje się tak w okresie stagnacji letniej. Jesienią podczas cyrkulacji, przy dobrym natlenieniu, wytrąca się z wody prawie cały zapas fosforu i osadza na dnie w postaci nierozpuszczalnego fosforanu żelazowego. Jest to zjawisko występujące cykliczne w zbiornikach z okresami przemieszania i stagnacji.
Fosfor rozpuszczony w wodzie pochodzi z lądu, jest wymywany z gleby (trudniej od azotu), oraz z rozkładu organizmów. Pierwiastek ten uwalnia się także z połączeń organicznych za pośrednictwem bakterii, oraz znacznie efektywniej w czasie rozkładu (autolizy) obumarłych organizmów wodnych. W tym ostatnim procesie czynny udział bierze enzym fosfataza, który odczepia fosfor z nukleoproteidów. Jest to proces szybki - już po 10 minutach z ciała nieżywych organizmów zaczyna wydzielać się fosfor nieorganiczny. Po 30 minutach od chwili śmierci już 12%, a po 24 godzinach 50-60% ogólnej ilości fosforu przechodzi w fosforany. Stąd też tempo cyrkulacji fosforu w wodzie jest szybsze niż azotu. Dlatego właśnie zapasy fosforu, mimo iż są mniejsze od azotowych, wystarczają na utrzymanie w normalnych warunkach procesów życiowych na odpowiednim poziomie.
Jak już wspomniano, w wodzie morskiej i w słodkiej fosforu jest mało - dziesiątki mg jonu PO4 w m3. Stosunek P:N kształtuje się na ogół jak 1:10. W Morzu Północnym w warstwach powierzchniowych notuje się przeciętnie około 20 mg fosforu w m3. Wraz z głębokością zawartość stopniowo rośnie. W jeziorach zawartość fosforu jest dość zróżnicowana. Na przebadanych 494 jeziorach w stanie Wisconsin (USA) w 20% jezior stwierdzono ślady fosforu lub jego brak, w 40% -2-3 mg/m3, w 18% - 4 mg, a w 6% - 6,7 mg w m3 wody. Dotyczy to oczywiście warstwy powierzchniowej w okresie stagnacji letniej. W jeziorach Pojezierza Bałtyckiego stosunki są podobne. Latem w epilimnionie zawartość fosforu wynosi przeciętnie 0-4 mg/m3, zaś w hypolimnionie 15-30 mg/m3. Natomiast podczas cyrkulacji zawartość jest wyrównana - wiosną 9-10 mg/m3, jesienią 6-8 mg/m3. W jeziorach meromiktycznych w monimolimnionie dochodzi do 800 mg/m3. Wahania sezonowe zawartości fosforu mają podobny przebieg jak azotu i uderzająca jest zależność między rozwojem fitoplanktonu a zasobami fosforu.
Materia organiczna
Pojęcie materii organicznej odnosi się do:
- postaci uorganizowanej jako składnika organizmów,
- zawiesiny obumarłych szczątków organicznych,
- związków organicznych w stanie koloidalnym lub rozpuszczonych w wodzie.
Materia ta może być naniesiona przez dopływy, opady atmosferyczne lub zmyta z brzegów, czyli pochodzić spoza zbiornika. Taki typ nosi nazwę materii organicznej allochtonicznej. Ta zaś, która powstaje w samym zbiorniku, jako wynik produkcji biologicznej, nazywa się materią organiczną autochtoniczną.
Różne są metody określania zawartości materii organicznej w wodzie. Jedna z nich to wyliczenie różnicy między pozostałością a wagą przed spaleniem odparowanej wody. Do zorientowania się w ilości materii organicznej rozpuszczonej w wodzie służy również oznaczenie tzw. utlenialności, czyli zużycia tlenu potrzebnego do utlenienia materii organicznej w określonych warunkach, lub. tzw. biochemicznego zapotrzebowania tlenu (BZT). Wyrażone jest ono poprzez ilość tlenu zużytego do utlenienia związków organicznych obecnych w wodzie, przy współudziale mikroorganizmów. Zawartość materii może być także wyrażana poprzez zawartość węgla organicznego lub azotu organicznego, względnie wzajemnego ich stosunku. W substancji autochtonicznej stosunek C:N wynosi 10-12:1, a w allochtonicznej 45-50:1. Materiał allochtoniczny jest ubogi w białka - najprawdopodobniej transport powoduje ich wyługowanie.
Stopień zawartości materii organicznej wskazuje na żyzność i produktywność biologiczną w zbiorniku wodnym. Oznaczenie składu tej materii jest bardzo trudne ze względu na jego niestabilność i dużą różnorodność związków tworzących ją. Rozpuszczoną materię organiczną stanowią produkty hydrolizy białek, węglowodanów i tłuszczów. Wiele z tych składników nadaje wodzie barwę brunatną np. kwasy humusowe.
Organizmy wodne mogą w różnym stopniu korzystać z materii organicznej, tak rozpuszczonej, jak i zawieszonej, czyli uorganizowanej. Jest ona przede wszystkim podstawą rozwoju bakterii i grzybów wodnych, gdyż dla nich stanowi źródło energii. Osmotycznie korzystają z niej miksotrofy, do których należą niektóre glony i drobne zwierzęta planktonowe. Sądzi się, że również ryby w pewnym zakresie mogą korzystać z rozpuszczonej materii organicznej. Duże ilości materii organicznej sprzyjają rozwojowi flory bakterii saprofitycznych będących doskonałym pożywieniem dla filtrujących zwierząt planktonowych - podstawy pokarmu wielu ryb.
Wpływ zlewni na chemizm wód
Zlewnia jest to obszar, na którym zbierają się i z którego spływają do zbiornika wodnego wszystkie wody powierzchniowe i podziemne. Zlewnia, jeśli dotyczy rzeki, często nazywana jest dorzeczem. Analogiczny obszar, odnoszący się do morza, nosi nazwę zlewiska. Ze zlewni do zbiornika, wskutek rozpuszczającego oddziaływania wody, przenoszone są różne jej składniki. Obszar, z którego zwiewane są wiatrem różnego typu zawiesiny, określa się nazwą zwiewni.
Rozpuszczanie, czyli wyciąganie z sieci krystalicznej minerału jakiegoś jonu, wymaga przezwyciężenia siły wiążącej go w tej sieci. Dokonać tego mogą naładowane dwubiegunowo cząsteczki wody. Gdy tzw. energia hydratacji - energia ładunku cząsteczki wody w procesie rozpuszczania - jest dostatecznie duża, następuje wyrwanie z sieci krystalicznej danego jonu i przeniesienie go do roztworu. Jony takie otoczone są otoczką z cząsteczek wody, czyli tzw. obwódką hydratacyjną. Proces ten w wodzie stojącej zostaje zahamowany z chwilą, gdy wyrówna się energia łączących się jonów i wytworzy się roztwór nasycony. Natomiast w wodzie bieżącej przybywają coraz to nowe cząsteczki wody, włączając się do procesu rozpuszczania podłoża.
Wymyte z podłoża substancje mogą pozostawać w postaci zdysocjowanej, w postaci koloidów, gdy składowe części rozpuszczonego związku są większe od 0,001 m, albo zawiesiny - gdy części składowe są większe od 0,1 m. Roztwór koloidalny bywa zabarwiony lub opalizujący (nieco mętnawy). W wodach naturalnych koloidy tworzy krzemionka i humus o ładunkach ujemnych oraz wodorotlenki żelaza i glinu o ładunkach dodatnich.
Skład chemiczny wód zależy od chemizmu zlewni, odnosi się to szczególnie do rzek. Zdolność do rozpuszczania skał przez wodę jest wzmacniana wskutek obecności w niej CO2, która staje się wtedy kwasem węglowym. W takiej formie rozpuszcza ona prawie wszystkie minerały skało- i glebotwórcze. Stopień rozpuszczania się skał jest zróżnicowany. Minerały w skali od najłatwiej do najtrudniej rozpuszczalnych można ułożyć następująco: sól kamienna> gips> kalcyt> dolomit> skaleniowce (nefelin, oliwin)> pirokseny i amfibole> serpentyn> plagioklazy bogate w anortyt (z Ca)> plagioklazy ubogie w anortyt (z Na)> ortoklaz> biotyt> kwarc> muskowit. Dwa ostatnie są praktycznie nierozpuszczalne w wodzie. Zdolność jonów do migracji do wody jest również zróżnicowana. Stosunkowo łatwo migrują do wody jony NO3, S, Cl, Br, w średnim stopniu jony Ca, Na, Mg, F, Sr, Zn, natomiast trudno większość metali i krzemionka. Tę różną zdolność do migracji pierwiastków określa stosunek zawartości danego pierwiastka w wodzie do jego całkowitej zawartości w podłożu. Jest to tzw. wartość klarkowa pierwiastka. Składa się na nią: rozpuszczalność związków, właściwości rozpuszczalnika oraz wielkość kompleksu sorpcyjnego podłoża zlewni i związana z tym łatwość przepływu wody w podłożu. Substancje ilaste i organiczne w podłożu będą stanowiły duży kompleks sorpcyjny.
W kształtowaniu się chemizmu wód biorą udział elementy naturalne, takie jak: podłoże, szata roślinna; oraz elementy zewnętrzne: klimat i działalność człowieka. Roślinność wpływa stabilizująco poprzez zatrzymywanie wody. Lasy zapobiegają wymywaniu z podłoża składników mineralnych, zmniejszają także erozję podłoża. Lasy iglaste mogą wpłynąć na zakwaszenie wody, gdyż z ich ściółki wymywane są fulwokwasy, będące składnikiem związków humusowych. Oddziaływanie klimatu związane jest z ilością opadów. Stopień wpływu wód opadowych uzależniony jest od specyfiki skał zlewni i wykształcenia szaty roślinnej. Z opadów część wody spływa bezpośrednio do zbiornika, część przesącza się tworząc wody gruntowe i wgłębne, które po jakimś czasie także docierają do zbiorników. W zależności od struktury skał proces natężenia infiltracji będzie różny. Szybko zachodzić będzie w skałach porowatych i przepuszczalnych, takich jak żwiry, piaski, zlepieńce i niektóre piaskowce. Trudniej w skałach porowatych i nieprzepuszczalnych, takich jak iły, gliny, margle i łupki ilaste. Praktycznie nie zachodzi w skałach nieporowatych i nieprzepuszczalnych - granitach, gnejsach, kwarcytach, łupkach krystalicznych i wapieniach.
Działalność człowieka w zlewni jest różnorodna. Nie ogranicza się tylko do zrzutu zanieczyszczeń komunalnych i przemysłowych. Dotyczy ona przede wszystkim gospodarki w zlewni, która winna być racjonalna. Niestety historia i współczesność dostarczają wiele przykładów skrajnej bezmyślności człowieka. Dokonywane na wielką skalę wyręby lasów prowadzą do zaniku jezior wskutek obniżenia lustra wód gruntowych. Wyręby o zasięgu mniejszym powodują przyspieszenie erozji, tym samym wzbogacenie mineralne zbiorników wodnych. Działalność rekultywacyjna na takich terenach nie jest prowadzona racjonalnie, np. zalesia się je iglastymi gatunkami drzew co prowadzi do zakwaszenia wody. Niewłaściwie wykonana melioracja jest klęską dla wód naturalnych, gdyż ułatwia przemieszczanie się składników biogennych. Stwierdzono, że stosunkowo łatwo wymywany jest azot w postaci azotanów, fosfor zaś trudniej ze względu na jego dużą sorbcję przez glebę oraz wytrącanie się z żelazem i glinem. Fosfor do wód dostaje się głównie ze ściekami komunalnymi i przemysłowymi.
Tak więc wpływ zlewni zależy od jej budowy geologicznej, charakteru gleby, typu roślinności, stopnia zagospodarowania rolniczego oraz typu zabudowy (miejska, wiejska, przemysłowa). Zlewnia wykształcona głównie na polach uprawnych dostarcza dużą ilość składników nawozowych: azotu, fosforu, potasu itp., które wpływają na zintensyfikowanie procesów produkcyjnych w wodzie. Obszary łąkowe w zależności od uprawy wpływają różnie. Uprawiane, dobrze nawożone, łąki będą tak jak pola przekazywały do zbiornika wody obfitujące w azot i fosfor. Łąki kwaśne i zatorfione, z występującą dużą ilością związków humusowych, powodować będą zakwaszenie wody zbiornika. Charakter oddziaływania zlewni leśnej zależy od rodzaju drzewostanu. Przewaga drzew iglastych w zlewni powoduje, że wody spływające z niej są kwaśne, a drzew liściastych, że woda jest obojętna lub zasadowa. Jednak bez względu na rodzaj drzew, ze zlewni leśnej zawsze spływają duże ilości substancji humusowych. Substancje te tworzą w wodzie koloidy, które strącają się pod wpływem rozpuszczalnych soli (szczególnie wapiennych) tworząc osad denny. Gdy wapnia jest niewiele, substancje humusowe utrzymują się długo w wodzie, nadając jej barwę brunatną.
Wody gruntowe są głównym źródłem wapnia, żelaza, manganu, glinu i krzemionki. Pierwiastki te, znalazłszy się w zbiorniku wodnym, podlegają dalszym reakcjom chemicznym i biologicznym, a w efekcie osadzają się na dnie. Wapń wytrąca się jako węglan wapnia w procesie biologicznego odwapniania wody. Również przy tej okazji wytrąca się częściowo fosfor w postaci fosforanu wapniowego. Rozpuszczone w wodzie sole żelazawe wytrącają się w postaci wodorotlenku żelazowego, przechodząc w obecności tlenu w środowisku alkalicznym z formy dwuwartościowej do nierozpuszczalnej trójwartościowej. Wodorotlenek ten osadzając się na dnie, gdzie są duże pokłady mułu organicznego, w obecności siarkowodoru tworzy czarny siarczek żelaza. Natomiast w wodach dobrze natlenionych z odczynem alkalicznym osadza się na dnie w postaci wodorotlenku żelaza, przechodząc w osad żelazisty, który z biegiem czasu tworzy pokłady rudy bagiennej lub jeziornej.
W zależności od charakteru przebiegu procesów chemicznych w wodzie tworzą się trzy typy osadów dennych. Sapropel jest osadem tworzącym się w wodach bardzo żyznych, w których przy dnie brak jest tlenu. Powstaje w wyniku procesów redukcyjnych i zawiera substancje organiczne nierozłożone. Osady te są zwykle czarne, kleisto-maziste, tłustawe. Gyttja (czytaj jüttia), powstaje w jeziorach eutroficznych. Przy jej tworzeniu grają rolę procesy redukcyjno-utleniające. Jest to szarozielona lub szara masa o zróżnicowanej ziarnistości, zawsze ze szczątkami organizmów lub odchodami. Dy są to osady powstające w ubogich jeziorach humusowych. Tworzą się w wyniku strącania się substancji humusowych pod wpływem soli, szczególnie wapiennych. Stanowią one ciemnobrunatną, zwykle gruboziarnistą, masę z brunatnymi wtrętami koloidów humusowych. Brak w nich szczątków i odchodów zwierzęcych.
W osadach zachodzą złożone procesy bakteryjnego rozkładu materii organicznej i tym samym powrotu do obiegu biologicznego związków nieorganicznych, potrzebnych roślinom do produkcji. Jest więc to osad biologicznie czynny, zasiedlony przez tzw. reducentów. Dolną granicę zasiedlenia określa obecność substancji pokarmowych oraz możliwość odprowadzania produktów przemiany materii, które zalegając zatruwają otoczenie i eliminują życie.
BIOLOGIA ORGANIZMÓW WODNYCH
Pływalność
Każde ciało znajdujące się w wodzie podlega prawom hydrostatyki, a wśród nich prawu Archimedesa. Gdyby takie ciało miało gęstość równą gęstości wody, to wyparte unosiłoby się przy powierzchni. Dlatego organizmy, aby żyć w toni wody, muszą mieć gęstość niewiele większą od gęstości wody. Powoduje to, że organizmy opadają. Szybkość tego opadania zgodnie z prawem Ostwalda zależy od ciężaru ciała, jego kształtu i lepkości wody, i wyraża się wzorem:
Równanie 3.
![]()
gdzie:
Vo - szybkość opadania
γk - opór kształtu,
ηw - lepkość wody
Δq - nadmiar ciężaru, który wylicza się według równania:
Równanie 4.
Δq = (qorg-qw)-worg
gdzie
qorg - gęstość organizmu,
qw - gęstość wody
worg - objętość organizmu,
.
Z powyższego wzoru wynika, że szybkość opadania jest tym większa, im większy jest ciężar organizmu, natomiast ogranicza ją opór wody, który zależy od kształtu organizmu i lepkości wody. Różnica pomiędzy ciężarem właściwym organizmu a ciężarem właściwym wody zależy w pewnym stopniu od temperatury (materia żywa zdolna jest do rozszerzania się pod wpływem ciepła) oraz innych czynników, np. stanu odżywienia organizmu. Czynniki te powodują ciągłe, choć nieznaczne, wahania gęstości ciała. Organizmy mają jednak możliwość przystosowania się i zmniejszenia tej różnicy poprzez pewne elementy budowy ciała, w skład których wchodzą substancje lżejsze od wody, takie jak gazy, śluz, czy tłuszcze. Zmniejszony może być również ciężar części szkieletowych.
Opór kształtu uzależniony jest od wielkości powierzchni właściwej oraz wielkości płaszczyzny rzutowania kształtu organizmu. Powierzchnia właściwa jest to stosunek powierzchni ciała do jego objętości. Najmniejszą powierzchnię właściwą ma kula, której prędkość opadania, zgodnie z prawem Stoksa, jest proporcjonalna do kwadratu promienia kuli. Niewiele organizmów ma kształt zbliżony do kuli, a wskutek obecności wyrostków, kolców, szczecin, listewek itp. ich powierzchnia właściwa zostaje zwiększona, co sprzyja wydłużaniu czasu ich pływalności. Służy temu również zwiększenie poziomej płaszczyzny rzutowania. Jeśli organizm o kształcie walca będzie opadał zawieszony pionowo, to rzut poziomy będzie równy powierzchni koła, czyli podstawy walca, gdy zaś będzie ułożony poziomo to kształt rzutu będzie prostokątem o powierzchni z pewnością większej od powierzchni koła. Tak więc opór wody znajdującej się pod walcem ułożonym poziomo będzie większy. Byłby on jeszcze większy, gdyby z boków walca rozmieszczone były listwy, powiększające rozmiary rzutu prostokąta. Odpowiednia lokalizacja środka ciężkości organizmów wodnych jest przystosowaniem mającym na celu utrzymanie najwłaściwszej pozycji w wodzie. Środek ciężkości organizmu umieszczony jest symetrycznie, tak aby umożliwiał łatwy powrót do najdogodniejszego położenia w wodzie. Służą do tego odpowiednio ustawione na ciele wyrostki, kolce oraz stateczniki, jak również ziarnistości rozmieszczone symetrycznie na końcach jednokomórkowego organizmu.
Lepkość wody jest czynnikiem fizycznym, który istotnie zależy od temperatury wody. Przy wzroście temperatury lepkość się zmniejsza, zmniejsza się i tarcie w wodzie, co powoduje zwiększenie prędkości opadania. Przystosowawczą odpowiedzią organizmów na to jest zwiększenie powierzchni ciała. Typowym przykładem na to jest zjawisko cyklomorfozy (patrz rozdz. Formacje ekologiczne). Jako przykład może posłużyć również porównanie organizmów planktonowych zamieszkujących ciepłe i zimne rejony Atlantyku. Bogactwo kształtów, polegające na wytworzeniu rozmaitych wyrostków i dzięki temu zwiększeniu powierzchni tarcia, jest typowe dla organizmów bytujących w strefie międzyzwrotnikowej. W wodach zimnych, gdzie lepkość jest większa, tego typu przystosowania są niepotrzebne i nieobecne.
Na swobodne unoszenie się organizmów w toni wodnej wpływ mają prądy turbulencyjne. Dzięki tym prądom oraz przystosowaniom biologicznym omawianym wyżej w wodzie może istnieć zawiesina, nazywana sestonem. W jego skład wchodzi zawiesina żywa - bioseston lub inaczej plankton, oraz nieżywa - abioseston. Seston nie może unosić się nieskończenie długo. Pomimo wszelkich przystosowań służących do unoszenia się w wodzie, aby być w niej zanurzonym, musi być od niej cięższy - tym samym kiedyś musi opaść na dno.
Ruch
Aktywne przemieszczanie się organizmów w wodzie polega na:
- ruchu amebowatym, czyli odkształcaniu plazmy,
- ruchu wiciowo-rzęskowym, czyli ruchu odpowiednich wyrostków ciała,
- ruchu za pośrednictwem aparatu mięśniowego.
Ruch wiciowo-rzęskowy jest typowy dla organizmów jednokomórkowych roślinnych i zwierzęcych. Wić wykonuje zwykle ruch drgający lub śrubowy, ciągnąc za sobą cały organizm. Większość organizmów ma tylko jedną wić. Spotyka się także kilka wici o jednakowej lub różnej długości. Występujące u niektórych organizmów rzęski są tworami podobnymi do wici, ale zwykle ustawionymi w rzędach. Wykonują one ruch falowy, powodujący odpychanie się od wody.
Ruch mięśniowy oparty jest na zasadzie skurczu i rozkurczu mięśni. U zwierząt pozbawionych sztywnego szkieletu ruch skurczowo-rozkurczowy mięśni powoduje skracanie lub wydłużanie ciała oraz jego zgięcia na boki. Najbardziej typowa odmiana ruchu mięśniowego to ten, w którym mięśnie oddziałują na zasadzie dźwigni na kończyny lub, w przypadku ryb, na ogon. U skorupiaków różne typy odnóży są swoim kształtem przystosowane do ruchu wiosełkowatego - spłaszczone lub zaopatrzone w rzędy wyrostków przypominają wiosła. Również u kręgowców odnóża służące do pływania przypominają wiosła.
Poruszanie się w wodzie jest trudniejsze niż na lądzie ze względu na większą gęstość środowiska wodnego od gęstości powietrza. Dlatego też w procesie tym istotną rolę odgrywa hydrodynamiczny, czyli opływowy, kształt ciała. Ogranicza on do minimum opór środowiska. Ciało opływowe ma kształt wrzeciona, w którym najbardziej rozszerzona część jest przesunięta do przodu i znajduje się w mniej więcej 1/3 długości. Przedni koniec jest zaokrąglony, a tylny wyciągnięty (rys. 23). Prócz kształtu ważnym elementem przy poruszaniu się w wodzie jest gładkość powierzchni ciała, zmniejszająca tarcie o wodę. Tak więc większość organizmów wodnych ma powierzchnię gładką lub czasami pokrytą śluzem. Natomiast te z organizmów, których ciało zaopatrzone jest w kolce lub innego typu wyrostki, wykazują przystosowanie polegające na utrzymaniu w tej nierównej powłoce ciała warstwy wody. Powoduje to, że podczas poruszania się organizmu trą się dwie warstwy wody, czyli ośrodki o tej samej gęstości, co wybitnie zmniejsza efekt tarcia.
Rys. 23. Przykłady opływowego kształtu ciała organizmów pływających (a-c) i poruszających się po dnie (d-j)
Innym przykładem ruchu organizmów wodnych jest zwykły ruch kroczący, typowy dla tych, które zasiedlają dno. Dla nich, wtedy gdy zamieszkują biotop mobilny, np. potok lub brzeg morza, opływowy kształt ciała jest również ważny.
Osmoregulacja i jonoregulacja
Przy omawianiu istoty oddziaływania czynników środowiska na organizm należy zwrócić uwagę na naturę środowiska wewnętrznego organizmu. Składnikiem podstawowym każdej komórki jest woda, która może stanowić ponad 95% jej masy. Tak więc upraszczając, każdy organizm jest grudką plazmy, oddzieloną od środowiska błoną półprzepuszczalną, tj. przepuszczalną dla wody, ale znacznie mniej przepuszczalną dla jonów. Efektem tego jest zróżnicowanie ciśnienia osmotycznego między wnętrzem komórki, lub organizmu, a otoczeniem. Stężenie soli jest na ogół większe w plazmie niż w środowisku zewnętrznym. Natomiast w wodzie tak silnie zasolonej jak morska, ciśnienie osmotyczne wnętrza komórki i otoczenia jest podobne. Zjawisko to nosi nazwę izotonii, a ciśnienie izotonicznego. U organizmów bytujących w środowisku słodkowodnym większe ciśnienie osmotyczne panuje we wnętrzu komórki niż w otoczeniu. To ciśnienie określa się nazwą hypertonicznego w stosunku do hypotonicznego w środowisku zewnętrznym. W takim przypadku, gdy ciśnienia te są różne, zachodzi proces zdążający do wyrównania ciśnienia.
Zjawisko osmozy, którego istotą jest wyrównywanie ciśnienia, będzie powodować stały napływ wody do wnętrza komórki. Proces ten mógłby w końcu doprowadzić do jej rozerwania. Aby temu zapobiec organizmy mogą wydalać nadmiar wody. To zjawisko nosi nazwę osmoregulacji. Przystosowania funkcjonalno-anatomiczne do osmoregulacji są różne i zależą od stopnia organizacji organizmu. U pierwotniaków funkcję osmoregulatora spełniają wodniczki tętniące. Wodniczki takie u form morskich występują rzadko, jeśli bytują one w układzie izotonicznym. U pozostałych grup bezkręgowców rolę taką spełniają narządy wydalnicze o specyficznej budowie zależnie od przynależności taksonomicznej. Natomiast u niektórych larw owadów (np. Chironomus) rolę taką częściowo spełnia końcowy odcinek jelita.
Zdolność regulacji ciśnienia osmotycznego jest zróżnicowana. Istnieje wiele organizmów, które mogą je zmieniać wraz z wahaniami stężenia soli w środowisku. Są to organizmy poikiloosmotyczne. Do nich zalicza się gąbki, jamochłony, pierścienice, mięczaki i niektóre skorupiaki. Inne, takie jak bardziej uorganizowane skorupiaki, larwy owadów wodnych oraz ryby, mają zdolność utrzymania mniej więcej stałego ciśnienia osmotycznego, niezależnie od środowiska. Te noszą nazwę organizmów homoioosmotycznych.
Jonoregulacja, w odróżnieniu od osmoregulacji (zjawiska odpowiedzialnego za gospodarkę wodną w organizmie), dotyczy wymiany elektrolitów między komórką a środowiskiem zewnętrznym. Każdy organizm wymaga dla zabezpieczenia swoich procesów życiowych pewnej równowagi jonowej. Wobec tego można mówić o bilansie wymiany jonowej, który został sformułowany przez Karandajewą:
Równanie 5.
![]()
gdzie:
Co - początkowe stężenie soli,
C1 - końcowe stężenie soli,
A - różnica między pozyskiwaniem a utratą soli na jednostkę ciężaru ciała,
S - osmotycznie czynna przestrzeń, wyliczana z różnicy między ciężarem masy świeżej i suchej,
P - ciężar początkowy
ΔP - zmiana ciężaru.
Zjawisko jonoregulacji jako proces biologiczny jest niezmiernie ważne w zbiornikach ubogich w sole mineralne, np. dystroficznych. Zachodzi w nich konieczność "rozważnej" gospodarki pulą jonów, które łatwo utracić, a znacznie trudniej pozyskać.
Oddychanie
Koncentrując się na aspekcie przystosowawczym procesu oddychania organizmów wodnych należy pamiętać o specyfice środowiska wodnego. W atmosferze zawartość tlenu jest prawie stała, w wodzie jest zaś uzależniona od warunków fizycznych, a przede wszystkim od temperatury. Zawartość tlenu podlega ponadto znacznym wahaniom dobowym spowodowanym różnym natężeniom procesu fotosyntezy w zależności od pory dnia. Na intensywność procesu oddychania wpływać może stopień zasolenia wody. Jest ono, według Thienemanna, łatwiejsze w wodzie słonej. Na przykład forma bałtycka skorupiaka Mysis relicta toleruje w wodzie słonej stężenie 1,6 cm3/l O2, natomiast w wodzie słodkiej nie wytrzymuje 4 cm3/l.
Tlen do oddychania bywa pobierany w rozmaity sposób. Rośliny i wiele zwierząt wodnych nie ma specjalnych narządów oddechowych i wymiana gazowa odbywa się całą powierzchnią ciała, drogą dyfuzji. Intensywność tego procesu zależy od powierzchni ciała danego organizmu stykającej się z wodą. Stąd też np. wywłócznik o silnie pierzastych liściach będzie mógł pobrać więcej tlenu w tym samym czasie niż morszczyn. Specjalnych narządów oddechowych brak jest u pierwotniaków, gąbek, jamochłonów, wirków, wrotków, niektórych pierścienic i drobnych skorupiaków. U wielu typowych mieszkańców wód w oddychaniu biorą udział różne narządy wewnętrzne. Meduzy i ukwiały pobierają tlen nie tylko poprzez zewnętrzną ścianę ciała, ale także powierzchnią jamy chłonąco-trawiącej. Strzykwy i larwy ważek wciągają wodę do odbytnicy i wykorzystują zawarty w niej tlen. Podobny typ oddychania jelitowego stwierdzono u wieloszczetów. Większość organizmów wodnych ma specjalnie wykształcone narządy umożliwiające oddychanie. Są to skrzela i skrzelotchawki.
Zwierzęta wodne wtórnie przystosowane do środowiska wodnego, np. wszystkie ssaki morskie, oddychają powietrzem atmosferycznym. Muszą więc od czasu do czasu wypływać na powierzchnię. Duża pojemność płuc i inne przystosowania pozwalają im na długie przebywanie pod wodą, np. wieloryb grenlandzki bez odnowienia powietrza w płucach może pływać pod wodą do 20 minut, a kaszalot nawet do półtorej godziny. Zimą, gdy powierzchnia wody jest zamarznięta, mieszkańcy rejonów biegunowych (np. foki) wykonują stale utrzymywane przeręble umożliwiające im kontakt z atmosferą. W tym samym celu wieloryby wędrują poza rejon lodowy. Owady wodne mają również zdolność korzystania z powietrza atmosferycznego. Niektóre z nich wynurzają się z wody i łapią pęcherzyk powietrza skrzydłami. Trzymając bańkę gazu między okrywami skrzydeł a powierzchnią odwłoka - tu mieszczą się przetchlinki - wciągają ją pod wodę. Wodne ślimaki płucodyszne (błotniarki i zatoczki) również oddychają powietrzem atmosferycznym, dlatego też występują w rejonach płytkowodnych, tuż przy brzegu.
Larwy komarów, np. widliszka, żyją w wodzie uczepione od spodu do błonki, tworzącej się w wyniku działania napięcia powierzchniowego na powierzchni drobnych zbiorników wodnych. Mają w ten sposób ułatwiony kontakt z powietrzem. Tę właściwość biologiczną komarów wykorzystano w metodzie zwalczania malarii za pomocą rozlewów ropy naftowej na powierzchni kałuż i bagien, co powoduje odcięcie dostępu powietrza. Innym sposobem zdobywania powietrza przez larwy owadów jest nakłuwanie liści roślin wodnych i wykorzystywanie tlenu zawartego w przestrzeniach międzykomórkowych. Tak więc różnorodność sposobów oddychania wpływa na przestrzenne rozmieszczanie organizmów w wodzie.
Zapotrzebowanie tlenowe organizmów wodnych jest bardzo zróżnicowane. Zależy przede wszystkim od potrzeb energetycznych, a związane jest z intensywnością ruchu. Zużycie tlenu przez formy planktonowe jest kilka razy mniejsze niż przez denne i kilkadziesiąt razy mniejsze niż przez formy nektonowe (tab. 4). Wymagania tlenowe zależą w znacznej mierze od temperatury wody. Zużycie tlenu stosuje się w pewnej mierze do reguły Vant Hoffa: szybkość reakcji chemicznych zwiększa się dwukrotnie o każde 10oC. W procesach fizjologicznych zużycie tlenu jest jeszcze intensywniejsze, np. łosoś pacyficzny w zależności od temperatury zużywa na 100 g wagi w ciągu godziny w około 8oC - 10,7 cm3, w 11,5oC - 21,4 cm3 , a w 19oC - 29,8 cm3 tlenu.
Tabela 4.
Zapotrzebowanie tlenowe na procesy respiracji (cm3O2/h.100 g biomasy)
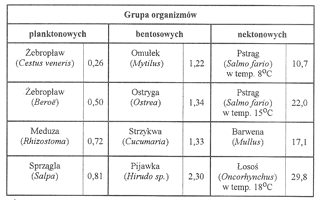
Źródło: Lityński A. Hydrobiologia ogólna. Warszawa, PWN 1952.
Zróżnicowana zawartość tlenu w wodzie ma istotny wpływ na rozwój i wzrost organizmów wodnych. Stwierdzono, że są pewne optymalne stężenia tlenu, przy których dany proces fizjologiczny przebiega najkorzystniej. W warunkach małej zawartości tlenu następuje ograniczenie wielu czynności, natomiast przekroczenie pewnego minimum powoduje obumieranie. Jedynie nieliczne gatunki mogą żyć w skrajnych warunkach tlenowych. Są nimi te, u których proces oddychania nie jest związany z tlenem rozpuszczonym w wodzie. Należą do nich bakterie siarkowe, żelaziste i inne czerpiące energię z anaerobowego rozkładu materii organicznej. Wiele gatunków wodnych ma zdolność oddychania śródcząsteczkowego polegającego na rozkładzie glikogenu zgromadzonego w organizmie. Jest to typowe dla zwierząt morskich żyjących na skraju strefy pływów, np. pąkli (Balanus). Przy niskiej wodzie mogą dłuższy czas żyć bez dostęu tlenu, szczelnie zamknięte płytkami swego szkieletu zewnętrznego. Niektóre z larw, np. larwa muchówki z rodzaju Chaoborus, żyją na dnie, czyli w środowisku, gdzie są śladowe ilości lub nawet brak tlenu. Wręcz zupełną odpornością na brak tlenu charakteryzują się wszelkie stadia przetrwalne, które po kilkumiesięcznym zimowym przebywaniu w mule dennym nie tracą zdolności życiowych.
Z ekologicznego punktu widzenia, ze względu na wymagania tlenowe wyróżnia się kilka grup organizmów wodnych. Są to euryoksybionty, czyli organizmy odporne na szerokie wahania zawartości tlenu w wodzie, oraz stenooksybionty, nie znoszące takich wahań. Jako przykłady pierwszej grupy mogą służyć takie skorupiaki planktonowe, jak Daphnia pulex, Diaptomus gracilis. W drugiej grupie wyróżnia się gatunki o dużych wymaganiach tlenowych, czyli polioksybionty (np. ryby łososiowate, a spośród skorupiaków Mysis relicta), oraz bytujące w środowisku beztlenowym lub ze śladowymi ilościami tlenu, czyli anoksybionty lub anaerobionty (np. rurecznik Tubifex tubifex żyjący w mule).
Uwzględniając zróżnicowanie wymagań tlenowych, Thienemann wyodrębnił dwie grupy organizmów przewodnich i w zależności od ich występowania w hypolimnionie latem wydzielił dwie grupy jezior. Są to zbiorniki tanytarsusowe, z przewagą w bentosie larw ochotek z grupy Tanytarsus, o dużych wymaganiach tlenowych. W hypolimnionie latem jeziora te nasycone są tlenem w ponad 50%. Obecne są w nich ryby łososiowate oraz skorupiaki reliktowe, takie jak Mysis relicta. Ze względu na charakter budżetu tlenowego są to jeziora oligotroficzne. Druga grupa obejmuje jeziora eutroficzne i według podziału Thienemanna są to zbiorniki chironomusowe, dla których charakterystyczny jest masowy rozwój larw ochotkowych z rodzaju Chironomus. W jeziorach tych w okresie stagnacji letniej zawartość tlenu przy dnie jest zawsze niższa od 50%, a w wielu przypadkach spada nawet do 0%. Wtedy gdy zawartość tlenu jest mniejsza niż około 5%, larwy ochotek znikają, a jedynym przedstawicielem makrofauny pozostaje larwa Chaoborus.
Odżywianie
Wyróżnia się dwa typy odżywiania: autotroficzne i heterotroficzne. Materia organiczna wytwarzana jest w procesie odżywiania autotroficznego w wyniku fotosyntezy, dzięki energii promienistej. Również w wyniku procesu chemosyntezy powstaje nowa materia organiczna z soli mineralnych przy użyciu energii wyzwolonej podczas rozkładu rozmaitych związków chemicznych.
Na intensywność fotosyntezy wpływ ma nie tylko ilość światła, jego zakres oraz powierzchnia asymilacyjna, o czym była mowa wcześniej, ale również ilość i charakter chemiczny dwutlenku węgla. Ze względu na różną postać CO2 wykorzystywaną w procesie fotosyntezy przez rośliny, Ruttner wydzielił trzy grupy roślin wodnych:
- Typ Fontinalis, obejmuje mchy wodne i krasnorosty; CO2 pobierany jest wyłącznie w stanie molekularnym, rozpuszczonym.
- Typ Elodea, obejmuje większość zanurzonych makrofitów; prócz rozpuszczonego CO2 jest pobierany także jon HCO3-, który rozszczepiany jest na CO2 i OH-, co prowadzić może nawet do zmiany pH środowiska. W typie tym pobieranie rozpuszczonego CO2 jest około 5 razy większe od wykorzystania HCO3-.
- Typ Scenedesmus, obejmuje większość planktonowych zielenic; głównym źródłem dwutlenku węgla jest HCO3-, który jest pobierany około 25 razy więcej niż CO2.
Cukry proste będące produktem fotosyntezy w dalszych etapach procesów autotroficznych podlegają przemianom na bardziej złożone substancje organiczne, takie jak białka, czy wreszcie tłuszcze. Do produkcji tego typu związków organicznych potrzebne są sole azotu, fosforu i siarki, a także niektórych metali. Od ich stężenia w wodzie zależy również ilość wyprodukowanej materii organicznej potrzebnej heterotrofom na podtrzymywanie procesów życiowych.
W sposobie heterotroficznego odżywiania się pozyskiwanie pokarmu zachodzi zwykle na zasadzie fagotrofii, czyli pochłaniania w całości cząstek pokarmowych. U niewielkiej grupy mało uorganizowanych zwierząt następuje osmotrofia (pasożytnicze pierwotniaki) lub tzw. pinocytoza, czyli wnikanie przez osłony ciała dużych cząstek organicznych. Wszystkie fagotrofy mają wykształcone specjalne narządy, które ułatwiają wprowadzanie pokarmu do przewodu pokarmowego. Ich kształt zależy od wielkości i zagęszczenia pokarmu, sposobu jego zdobywania i od jego jakości.
Ze względu na wielkość pokarmu wyodrębnia się dwie grupy zwierząt: makrofagi, pobierające stosunkowo duży pokarm, niekiedy ich własnych rozmiarów, oraz mikrofagi, żywiące się pokarmem drobnym, o wiele od nich mniejszym.
Makrofagi to przede wszystkim drapieżcy oraz niektórzy roślinożercy żywiący się dużymi roślinami. U drapieżców istotnym elementem przystosowawczym są narządy chwytne, zwykle rozmieszczone niedaleko otworu gębowego. Są to przeważnie czułki, macki lub silne zęby, które służą do schwytania ofiary, przytrzymania jej i ćwiartowania. Również makrofagi-roślinożercy mają podobne narządy, ale odpowiednio zmodyfikowane.
Mikrofagi wymagają innych przystosowań. Pokarm rozproszony w postaci drobnej zawiesiny, aby mógł być wykorzystany jako pożywienie, musi zostać zagęszczony i pobrany. Odbywa się to dwoma sposobami, albo poprzez sedymentację, albo filtrację, czyli odsączanie.
Sedymentacja jako proces zdobywania pokarmu zachodzi z większym lub mniejszym udziałem organizmu. Pierwszy typ to tzw. sedymentacja czynna - zwierzę wywołuje prąd wody w postaci wiru, w wyniku czego cząstki pokarmu napędzane są w okolice otworu pokarmowego i tam się osadzają. Jest to typowy sposób odżywiania się wrotków, u których sedymentacja czynna zachodzi dzięki obecności aparatu wrotnego, oraz orzęsków, które wywołują wiry wody za pomocą rzęsek (rys. 25).
Rys. 24. Funkcjonowanie aparatu wrotnego w procesie sedymentacji czynnej.
Drugi typ sedymentacji to sedymentacja bierna. Polega ona na wytwarzaniu przez zwierzęta różnego typu sit, zastawek itp. w formie narzędzi łownych. Ustawiane są one zwykle w wodzie ruchomej, np. płynącej. Wskutek przemieszczania się wody na sitach osadza się zawiesina, która jest zjadana razem z zastawionymi uprzednio sitami (rys. 25).
Rys. 25. Przykłady sit chruścików wykorzystywanych w procesie sedymentacji biernej.
Filtracja jest sposobem czynnego zdobywania pożywienia. W tym celu wykształcone są specjalne narządy, najbardziej charakterystyczne u skorupiaków planktonowych. U wioślarek aparat filtracyjny tworzą spłaszczone odnóża zaopatrzone w wyrostki ze szczecinami, w postaci jakby grzebienia. Przy odpowiednim ustawieniu odnóży tworzy się układ sit zatrzymujących cząsteczki pokarmowe, które przesuwane są następnie w kierunku otworu gębowego. U rozwielitek, dzięki odpowiedniemu ustawieniu odnóży, tworzą się komory, które są na przemian napełniane wodą i opróżniane. Typowymi filtratorami morskimi są wieloryby fiszbinowe. Nabierają one w ogromną paszczę wodę, która następnie wypychana językiem przesącza się przez sita fiszbinów. Osadzony w nich kryl jest następnie zjadany.
Mikrofagami są również małże pozyskujące pokarm z wody dostającej się syfonem wpustowym do jamy płaszczowej. Równocześnie podczas obmywania skrzeli następuje pobranie do oddychania tlenu zawartego w wodzie. Małże należy więc zaliczyć do sedymentatorów. Ślimaki, które żywią się peryfitonem, czyli osiadłymi na różnych przedmiotach drobnymi glonami i zwierzętami nazywa się skrobaczami. Nazwę zawdzięczają specjalnie wykształconemu narządowi gębowemu - tarce, którą zeskrobują porośla znacząc tym samym przebytą drogę. Tak więc u ślimaków i innych podobnie żywiących się zwierząt pozyskiwanie pokarmu związane jest z ich czynnym przemieszczaniem się.
Specyficzną grupą mikrofagów są masojady, które pobierają bardzo duże ilości masy pokarmowej, z czego zaledwie niewielka część nadaje się na pożywienie. Przykładem są zwierzęta żyjące na dnie i żywiące się mułem organicznym tam osadzonym. Do tej grupy należą przede wszystkim skąposzczety i wieloszczety. Rureczniki żerują w osadach na głębokości do 3 m, zaś larwy ochotek żywią się mikroglonami żyjącymi na powierzchni mułu.
Pobrany pokarm musi pokryć wydatki energetyczne na podstawowe procesy fizjologiczne organizmu. Dlatego też każdy organizm potrzebuje w określonym czasie odpowiedniej ilości pokarmu, czyli racji pokarmowej. Przewód pokarmowy może być napełniony kilka razy w ciągu doby, zależy to od pojemności przewodu oraz rodzaju pożywienia. Ciężar napełnionego jednorazowo przewodu pokarmowego wyrażony w procencie masy ciała zwierzęcia nazywa się wskaźnikiem nasycenia. Z wartości tego wskaźnika (n) i czasu potrzebnego na przejście jednorazowego napełnienia przez przewód (t) można wyliczyć dobową rację pokarmową
Równanie 6.
R=24n/t.
Wielkość racji pokarmowej świadczy o intensywności odżywiania. U filtratorów tempo filtracji zależy od takich czynników zewnętrznych, jak: temperatura, światło oraz zagęszczenie pokarmu w wodzie. Racja pokarmowa jest zależna od wielkości organizmu, a tym samym jego ciężaru. Zwykle jest duża u mniejszych, a spada u większych. Zmiana racji pokarmowej może być także spowodowana raptownym przejściem na inny rodzaj pokarmu. Skoro intensywność odżywiania zależy od warunków środowiskowych, a przede wszystkim temperatury oraz nasycenia wody tlenem, to tak jak te czynniki, wykazuje ona rytmiczność dobową i sezonową.
Wybiórczość pokarmowa jest to zdolność do zróżnicowanego wykorzystania masy białkowej znajdującej się w zasięgu zwierzęcia. Określić ją można za pomocą wskaźnika Szorygina:
Równanie 7.
I = b/a
gdzie:
a - procentowa zawartość danego składnika pokarmowego w środowisku (np. w sestonie),
b - procentowa zawartość tego składnika w przewodzie pokarmowym (np. żołądku drapieżnika).
Gdy w planktonie zjadany gatunek np. wioślarki stanowi 50%, a w żołądku ryby zaledwie 5%, wówczas wskaźnik I = 0,1. Jest on mały, więc można sądzić, że albo ryba ta specjalnie nie wybiera rozwielitki, albo rozwielitka ma zdolność jej unikania. Ta dwuznaczność została wyeliminowana dzięki wskaźnikom Konstantinowa:
Równanie 8. Wskaźnik wybiórczości Konstantinowa.
I = (a-b)/a
Równanie 9. Wskaźnik unikania Konstantinowa.
I' = (a-b)/b
Wskaźnik wybiórczości przyjmuje wartości od 0 do 1, a unikania od 1 do nieskończoności.
Biorąc za kryterium wybiórczość pokarmową wyróżnić można zwierzęta wszystkożerne - polifagi lub euryfagi, oraz żerujące na jednym typie pokarmu - monofagi lub stenofagi. Do tej drugiej gupy zalicza się organizmy roślinożerne (fitofagi) lub zwierzęcożerne (zoofagi), które dalej można zróżnicować, np. z grupy ryb planktonożernych niektóre żywią się jedynie wioślarkami. Wybiórczość pokarmowa jest cechą wrodzoną, może się ona jednak zmieniać w różnych okresach roku i także w różnych fazach cyklu rozwojowego.
Specyficzny mechanizm odżywiania występuje u bakterii, które z tego względu dzielone są na trzy grupy: fotoautotroficzne, chemoautotroficzne i heterotroficzne. Dwie pierwsze grupy to bakterie samożywne, przy czym fotoautotrofy przyswajają dwutlenek węgla na świetle, a chemoautotrofom światło w tym procesie nie jest niezbędne.
Bakterie fotoautotroficzne mają barwniki do przeprowadzenia fotosyntezy, która różni się od fotosyntezy roślin zielonych tym, że wodór nie pochodzi z rozkładu wody a z innych zredukowanych związków np. z H2S, kwasów organicznych lub innych związków organicznych. Barwniki asymilacyjne w tej grupie bakterii to:
- bakteriowirydyna - barwnik zielony, podobny do chlorofilu a, o niejasnej budowie chemicznej;
- bakteriopurpuryna - barwnik czerwony, zbliżony do karotenoidów;
- bakteriochlorofil - barwnik zielony, zbliżony do chlorofilu a, o sumarycznym wzorze C55H74O6N4Mg (wzór chlorofilu a - C55H72O5N4Mg).
Wśród fotoautotrofów wyróżnia się bakterie zielone (Chlorobacteriaceae) z bakteriowirydyną i bakterie purpurowe (Thiorhodaceae) z bakteriochlorofilem lub bakteriopurpuryną. Wymagają one mineralnych dostawców wodoru do procesu fotosyntezy, która przebiega zgodnie z reakcją:
CO2 + 2H2S ------ energia świetlna (+ 30,2 cal) ------> (CH2O) + H2O + 2S
Istnieje jeszcze grupa bakterii purpurowych bezsiarkowych (Athiorhodaceae), które wykorzystują wodór z kwasów organicznych. Schematycznie proces fotosyntezy u bakterii fotoautotroficznych (siarkowych i bezsiarkowych) przebiega następująco:
CO2 + 2H2A ------ energia świetlna------> (CH2O) + H2O + 2A
gdzie:
- u bakterii siarkowych A = S (siarka)
- u bakterii bezsiarkowych A = R (rodnik organiczny).
W tej grupie bakterii intensywność fotosyntezy zależy od natężenia światła, stężenia CO2, temperatury wody i stężenia dostawcy wodoru.
Bakterie chemoautotroficzne do rozszczepienia CO2 wykorzystują energię procesów egzotermicznych, które same stymulują. Ta uwolniona energia spożytkowana jest do asymilacji CO2 przez bakterie; jest to proces endotermiczny i przebiega według schematu:
CO2 + H2O ------> (CH2O) + O2 - 113 kcal
Ponieważ energia ta pochodzi z reakcji chemicznych, przyjęto określać ten proces nazwą chemosynteza. W zależności od związków, które podlegają utlenianiu, bakterie nazwano siarkowymi, żelazowymi, manganowymi, wodorowymi, metanowymi itp.
Bakterie siarkowe to formy na ogół nitkowate, bezbarwne, z drobnymi ziarnkami siarki, która tworzy się w wyniku utleniania siarkowodoru w myśl reakcji:
2H2S + O2 ------> 2H2O + 2S + 118 kcal
Gdy wyczerpie się w środowisku siarkowodór to siarka zmagazynowana w komórkach utlenia się tworząc kwas siarkowy, który w obecności węglanu wapnia ulega zobojętnieniu przechodząc w siarczan wapnia. Tak więc chemoautotroficzne bakterie siarkowe usuwają ze środowiska trujący siarkowodór i wytwarzają siarczany potrzebne roślinom do budowy białek.
Bakterie żelazowe i manganowe prowadzą do utlenienia rozpuszczalnych węglanów żelaza i manganu, tworząc nierozpuszczalne wodorotlenki, według reakcji:
2Fe(HCO3)2 + 1/2O2 + H2O ------> 2 Fe(OH)3 + 4CO2
Wytrącający się wokół nitkowatych form bakterii żelazowych, a potem opadający na dno wodorotlenek żelaza tworzy w odpowiednich warunkach pokłady rudy żelaza.
Bakterie manganowe są mało znane; proces utleniania jest taki sam jak w wypadku żelaza. Bakterie wodorowe utleniają wodór:
2H2 + O2 ------> 2H2O + 138 kcal
zaś metanowe - metan:
CH4 + 2O2 ------> CO2 + 2H2O + 195 kcal
a CO-tlenkowe - tlenek węgla:
2CO + O2 ------> 2CO2 + 148 kcal
Bakterie nitryfikacyjne wykorzystują energię uwalnianą przy utlenianiu amoniaku do azotynów, a dalej azotanów. Bakterie azotynowe (Nitrosomonas) utleniają amoniak tworząc kwas azotawy:
2NH4OH + 3O2 ------> 2HNO2 + 4H2O + 132 kcal
a bakterie azotanowe (Nitrobacter) utleniają kwas azotawy tworząc kwas azotowy:
2HNO2 + O2 ------> 2HNO3 + 48,4 kcal
Procesem odwrotnym do nitryfikacji jest denitryfikacja, polegająca na uwalnianiu wolnego azotu z jego powiązań mineralnych (redukcja azotanów poprzez azotyny do wolnego azotu). Zachodzi ona z udziałem odpowiednich gatunków bakterii, w warunkach beztlenowych lub przy niskiej zawartości tlenu, w podwyższonej temperaturze i przy obecności dużych ilości węglowodanów. Z powodu zwiększania zapasów azotu przyswajalnego przez rośliny w wodach istotną rolę odgrywają bakterie i glony wiążące wolny azot.
Bakterie heterotroficzne oraz grzyby odżywiają się materią organiczną. Przyswajają ją i wykorzystują do budowy własnego ciała. Energia wyzwalana podczas jej rozkładu jest również zużywana w procesach życiowych. W tym celu mikroorganizmy rozkładają najpierw białka, cukry i skrobię, a potem trudniej rozkładające się substancje, takie jak celuloza, pektyny, ligniny. Końcowymi produktami rozkładu są także gazy, np. dwutlenek węgla, amoniak, wodór, siarkowodór, metan, oraz sole mineralne i organiczne związki humusowe. Całkowita mineralizacja materii organicznej może przebiegać jedynie w obecności tlenu, czyli w wodach natlenionych, natomiast w warunkach beztlenowych rozkład jest niecałkowity. Tak więc bakterie heterotroficzne i grzyby mogą wykorzystywać węgiel potrzebny do budowy własnego ciała jedynie z gotowej, odpowiednio rozłożonej materii organicznej. W procesach rozkładu biorą udział specyficzne enzymy, które mogą działać wewnątrz komórki (endoenzymy) lub są wydalane do środowiska i stymulują przemiany biochemiczne poza komórką (ektoenzymy). W pierwszym etapie następuje hydroliza materii organicznej pod wpływem działania ektoenzymów. Produkty tej fazy rozkładu pobierane są przez bakterie. W komórce bakterii w wyniku działania endoenzymów następuje dalszy ich rozkład. Część z nich jest wykorzystywana jako materiał budulcowy własnego ciała, część wraca do środowiska i może być wykorzystywana przez inne formy bakteryjne. Za poszczególne etapy rozkładu odpowiednich związków odpowiedzialne są specyficzne enzymy.
Rozmnażanie i rozwój
Wyróżnia się dwie formy rozrodu: płciowy i bezpłciowy, określany także jako wegetatywny. Rozród płciowy polega na produkowaniu przez organizm specjalnych komórek rozrodczych - komórek płciowych. Komórki te zwane gametami (jaja i plemniki) po połączeniu dają zygotę, która poprzez szereg przeobrażeń w cyklu rozwojowym daje postać dorosłą, zdolną znowu do produkcji gamet. Rozród wegetatywny polega na rozdzielaniu się organizmu macierzystego, np. przez podział na dwie części, które wzrastając uzupełniają pozostałą część, lub na oddzieleniu się części, z których wyrasta organizm dorosły, np. przez pączkowanie. Rozród bezpłciowy jako szczególny przypadek rozmnażania wegetatywnego polega na wytwarzaniu specjalnych komórek typu pływek. Tworzą się one bez uprzedniego podziału materiału genetycznego, jak dzieje się to w przypadku gamet. Specyficzny przypadek rozmnażania wegetatywnego to różnego typu spory, czyli tzw. komórki przetrwalne występujące dość często u roślin - szczególnie glonów planktonowych. U zwierząt polega to na ocystowaniu się całych organizmów. Po przebyciu niekorzystnych warunków środowiskowych następuje powrót do normalnych funkcji życiowych.
Woda, ze względu na pierwotny ewolucyjnie charakter tego środowiska, jest dogodnym środowiskiem dla różnego typu rozrodu. Także organizmy lądowe do rozmnażania i rozwoju potrzebują środowiska płynnego. W procesie rozrodu i rozwoju organizmy wodne wykazują różnorodne przystosowania do środowiska. Jest to wyraźnie widoczne u glonów rozmnażających się wegetatywnie przez fragmentację plech i podział komórki oraz poprzez wytwarzanie różnego typu zarodników. Ponadto prawie u wszystkich glonów wytwarzane są komórki płciowe, czyli gamety. Rozród wegetatywny u naczyniowych roślin wodnych jest zjawiskiem pospolitym dzięki łatwemu rozrastaniu się kłączy i rozłogów, np. u grzybieni czy rdestnic, lub fragmentacji, gdy z części rośliny powstają nowe osobniki jak np. u rogatka lub moczarki. Także w rozrodzie płciowym spotyka się wiele przystosowań wynikających z przebywania w wodzie. Są rośliny zapylane przez ślimaki wodne lub chrząszcze. Pyłki niektórych roślin są biernie unoszone w wodzie, dopóki nie trafią na kwiaty żeńskie.
Rozmnażanie wegetatywne u zwierząt jest typowe dla form prymitywnych. Przez podział rozmnażają się pierwotniaki i jamochłony. U tych ostatnich częstym zjawiskiem jest pączkowanie. Jednak podstawowym sposobem rozmnażania dla zwierząt jest rozród płciowy, związany z wymianą materiału genetycznego. Zapłodnienie zwykle jest zewnętrzne, szczególnie powszechne w wodzie morskiej ze względu na izotonię ciśnienia osmotycznego. Również z tego powodu w morzu powszechne są jaja pelagiczne, u zwierząt słodkowodnych będących raczej rzadkością. Jaja w zbiornikach śródlądowych opadają na dno przyczepiając się do podłoża albo są przez samicę składane na dnie lub przyczepione do roślin wodnych. Służą temu osłony jaj zaopatrzone w rozmaitego rodzaju wyrostki, haczyki i inne podobne twory ułatwiające przyczepianie się do substratu. Dość powszechnym zjawiskiem u organizmów wodnych jest dzieworództwo, czyli rozwój partenogenetyczny. Partenogeneza może występować przez wiele kolejnych pokoleń jako przeważająca forma rozrodu.
W rozwoju pozazarodkowym występuje larwa, bardzo powszechna u form morskich. Prawie wszystkie gatunki morskie przechodzą w rozwoju stadium larwy. U większości gatunków słodkowodnych stadium larwalne jest uproszczone lub w ogóle go brak. U wielu grup owadów wodnych przeobrażenie jest niezupełne i wówczas w pierwszym okresie rozwoju występuje tzw. larwula, która powoli upodabnia się budową ciała do postaci dorosłej (imago). W zależności od trybu życia larwy owadów wodnych wykazują daleko idące przystosowania w budowie. Skrajnym przykładem są larwy chruścików, które budują różnorodne domki ochronne w zależności od typu środowiska i dostępnego materiału. Każdy gatunek ma typowy czas trwania okresu larwalnego. Mają nań jednak wpływ czynniki ekologiczne, w tym przede wszystkim temperatura.
Formacje ekologiczne
Formacja ekologiczna obejmuje organizmy i ich stadia rozwojowe charakteryzujące się podobieństwem w specyfice biologicznych przystosowań do zasiedlanego środowiska. Są to obszerne grupy gatunków zasiedlające toń wody względnie dno zbiornika. Wyróżnia się pięć formacji ekologicznych: dwie bytujące w toni wodnej - plankton i nekton, dwie związane z warstwą styku woda-atmosfera - neuston i pleuston, oraz jedną zasiedlającą dno - bentos.
Plankton
Nazwę planktonu wprowadził do nauki Hensen w 1887 roku. Pochodzi ona od słowa greckiego planktos (tzn. błąkający się) i obejmuje organizmy roślinne i zwierzęce unoszące się swobodnie w wodzie, przeważnie o niewielkich rozmiarach ciała. Organizmy te albo nie mają wcale narządów ruchu, albo narządy te są na tyle słabe, że nie pozwalają na przeciwstawianie się silniejszym ruchom wody i uniemożliwiają poruszanie się pod prąd.
Ze względu na skład dzieli się plankton na dwie zasadnicze grupy: fitoplankton - plankton roślinny i zooplankton - plankton zwierzęcy. Niektórzy autorzy wyróżniają jeszcze jedną grupę obejmującą bakterie planktonowe, czyli bakterioplankton.
W zależności od wielkości organizmów wyróżnia się w planktonie kilka frakcji:
- megaloplankton, który obejmuje duże formy zwierzęce, o wielkości powyżej 1 cm; jest typowy dla mórz, w wodach śródlądowych trafia się wyjątkowo; do tej frakcji zalicza się między innymi duże meduzy;
- makroplankton, o wielkości 1-10 mm, składa się głównie z większych skorupiaków;
- mezoplankton, o wielkości od 500 mm do 1 mm, obejmuje mniejsze formy skorupiaków, oraz niektóre glony kolonijne;
- mikroplankton, o wymiarach w granicach 60-500 mm, należy tu większość glonów planktonowych oraz drobniejsze zwierzęta, w tym przede wszystkim wrotki;
- nannoplankton obejmuje najdrobniejsze wiciowce, glony i większe bakterie w zakresie wielkości 5-60 mm;
- ultraplankton jest to właściwie plankton bakteryjny o wymiarach mniejszych niż 5 mm.,
Ze względu na typ zbiornika, w którym plankton występuje, wyróżnia się kilka jego kategorii:
- haliplankton - plankton mórz;
- hyphalmiroplankton - plankton słonawych ujść rzecznych lub mórz o słabym zasoleniu;
- salinoplankton - plankton śródlądowych wód słonych;
- limnoplankton - plankton wód słodkich.
Limnoplankton dzieli się z kolei na: eulimnoplankton (z greckiego limne jezioro) - plankton jezior, heleoplankton (gr. helos - moczar, staw) -plankton stawów, potamoplankton (gr. potamos - rzeka) - plankton rzek, krenoplankton (gr. krene - źródło) - plankton źródeł, telmatoplankton (gr. telma - kałuża) - plankton kałuż i innych krótkotrwałych zbiorników.
Pod względem stopnia związania z formacją planktonu wyodrębnia się trzy grupy organizmów:
- Euplankton, czyli plankton właściwy obejmuje te gatunki, które prawie cały cykl życiowy spędzają zawieszone w toni wodnej. Jedynie okres spoczynkowy w postaci cyst, akinet itp. mogą przechodzić w szlamie dna.
- Meroplankton są to organizmy, które zasadniczy cykl swojego życia spędzają jako osiadłe, a jedynie w pewnym stadium tego cyklu prowadzą życie planktonowe. Odnosi się to do różnego typu zoospor glonów oraz ruchliwych stadiów (stadium larwalne) zwierząt osiadłych lub prowadzących denny tryb życia. Tu również zalicza się pelagiczną ikrę ryb.
- Tychoplankton obejmuje te organizmy, które przypadkowo znalazły się w planktonie, a normalnie żyją na dnie lub jako przytwierdzone do innych. Wykształca się on wskutek ruchu wody w małych zbiornikach wodnych lub w rzekach. Organizmy te czasowo występują w toni wodnej, po ustaniu przyczyny powodującej mieszanie wracają do właściwego im środowiska.
Występowanie planktonu w zbiornikach naturalnych nie jest jednorodne. Czynniki wpływające na jakość i ilość planktonu oraz na jego rozmieszczenie w wodach są różne. Można je ująć w dwie grupy. Pierwsza grupa to czynniki środowiskowe oddziałujące mechanicznie na organizmy. Jest to położenie zbiornika wodnego w stosunku do działania wiatrów, tym samym stopień jego odsłonięcia. Następnie zmienność stanów wody, ruchu wody w samym zbiorniku (wszelkiego typu prądy). Obecność pokrywy lodowej i wykształcenie uwarstwienia termicznego oraz stosunki gęstościowe w zbiorniku. Druga grupa czynników, to te które wpływają na biologię organizmów planktonowych, czyli o oddziaływaniu ekologicznym. Są nimi temperatura, światło, biogeny oraz materia organiczna, a w wielu przypadkach ścieki organiczne o różnym stopniu mineralizacji.
Fitoplankton składa się z sinic oraz z kilku grup glonów: euglenin, tobołków, złotowiciowców, okrzemek, różnowiciowców oraz obszernej grupy zielenic. W fitoplanktonie morskim obecne są jedynie niektóre grupy, a przede wszystkim okrzemki, z tobołków bruzdnice, zaś spośród wiciowców - wapienne i krzemionkowe. W związku ze specyfiką ekologiczną poszczególnych gatunków, a nawet całych grup fitoplanktonu, notuje się wyraźne zróżnicowanie w dominantach w zależności od typu zbiornika wodnego. W przeżyźnionych i dobrze nagrzanych stawach dominują sinice, w głębokich i ubogich jeziorach liczne są bruzdnice, w bogatszych w sole biogenne dominują okrzemki, zaś w zbiornikach humusowych przeważają zielenice-desmidie. To wyraźne przywiązanie danej grupy fitoplanktonu do typu siedliska było podstawą do wydzielenia przez Hutchinsona kilkunastu zespołów ekologicznych w planktonie roślinnym. Właściwość tą wykorzystali również Thunmark i Nygaard, wprowadzając system indeksów fitoplanktonowych, w których poprzez wzajemny stosunek liczby gatunków poszczególnych grup taksonomicznych fitoplanktonu ocenić można stopień produktywności zbiornika (tab.5). Im wartość wskaźnika jest większa, tym zbiornik jest żyźniejszy.
Tabela 5
Współczynniki fitoplanktonowe Nygaarda
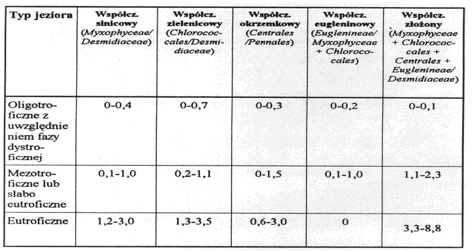
Źródło: Półtoracka J. Współczynniki fitoplanktonowe jako metoda określania stopnia troficzności środowisk wodnych. Ekologia Polska 1969, ser. B, 15/2.
Zooplankton jest bardziej zróżnicowany biologicznie aniżeli fitoplankton. W jego skład wchodzi kilka grup taksonomicznych: pierwotniaki, wrotki, jamochłony, żebropławy, pierścienice, strzałki i stawonogi oraz stadia larwalne mięczaków i szkarłupni, a także proste formy kręgowców oraz ikra i stadia młodociane ryb. Dla zooplanktonu słodkowodnego typowe są heterotroficzne pierwotniaki, wrotki oraz skorupiaki.
Odżywianie zooplanktonu odbywa się drogą sedymentacji czynnej lub filtracji. W niezbyt dużej liczbie gatunków notuje się drapieżnictwo. W rozwoju zooplanktonu cechą charakterystyczną jest obecność kilku stadiów rozwojowych, następujących po sobie. Klasycznym przykładem tego typu rozwoju są widłonogi (Copepoda) (rys. 26). Zooplankton stanowi bazę pokarmową dla wielu grup zwierząt, stąd też jego duża intensywność namnażania się jest ważnym elementem biologicznym. Intensywności tej służy kilka przystosowań: krótki okres rozwoju zarodkowego jaj, skrócenie rozwoju postembrionalnego, szybkie następstwo pokoleń (kilka w jednym roku)), zdolność rozrodu partenogenetycznego.
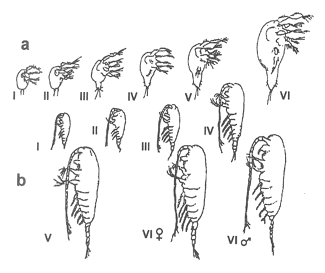
Rys. 26. Stadia rozwojowe widłonogów a - nauplialne b - kopepoditowe
Liczebność poszczególnych gatunków w cyklu rocznym nie jest równomierna. W zooplanktonie wyróżnia się gatunki całoroczne, które jednak wykazują maksimum rozwoju w określonych sezonach (np. Keratella quadrata ma szczyt liczebności w maju i czerwcu a K. cochlearis w lipcu i sierpniu). Ponadto są gatunki występujące sezonowo, a mianowicie zimowo-wiosennie, oraz latem i jesienią.
Rozmieszczenie pionowe zooplanktonu w zbiorniku wodnym kształtowane jest przez kompleks czynników środowiskowych, z których największy wpływ wywiera temperatura. Niezmiernie ważnym elementem rozmieszczenia pionowego zooplanktonu są jego wędrówki pionowe. W ich wyniku organizmy przemieszczają się przez ekologicznie zróżnicowane warstwy, w których są różne stosunki oświetlenia, temperatury, natlenienia i zasobów pokarmowych. Wędrówki te wykazują zadziwiającą regularność w cyklu dobowym i dlatego określa się je mianem wędrówek dobowych. Niektóre gatunki fitoplanktonu charakteryzują się pewnym specyficznym typem wędrówek dobowych. Dotyczy to glonów ruchomych, wiciowcowych. Wędrówki te polegają na przemieszczaniu się w dzień do rejonów o najlepszych warunkach świetlnych i mają charakter fototaksacji dodatniej. Zjawisko to notowane jest u bruzdnic i wiciowcowych zielenic. Stwierdzono, że pozytywna reakcja związana jest ze ściśle określonym stopniem natężenia światła, typowym dla danego gatunku. Bardzo mocne naświetlenie wywołuje reakcję negatywną.
Pionowe wędrówki dobowe zwierząt najlepiej zostały poznane u skorupiaków. Ogólnie schemat cyklu wędrówki dobowej obejmuje cztery fazy:
- energiczne przemieszczanie się w dół po wschodzie słońca;
- zaleganie na poziomie najniższym dla danego gatunku w ciągu godzin rannych i południowych;
- stopniowe wydźwiganie się ku górze w godzinach popołudniowych, z osiągniętym maksimum pod powierzchnią wieczorem;
- stopniowe opuszczanie się w dół nocą, co robi wrażenie równomiernego rozmieszczenia w pionie.
Liczne badania nad przyczynami wędrówek pionowych nie dały jednoznacznej odpowiedzi. Oczywiste jest ich zsynchronizowanie z ruchem słońca na niebie. Nie bez znaczenia jest tu również zależność troficzna - przesuwanie się zwierząt ku górze następuje po południu, czyli wtedy, gdy efekt produkcji biomasy roślinnej jest najpełniejszy w trakcie doby.
Innym zjawiskiem biologicznym dotyczącym tak fito- jak i zooplanktonu jest cyklomorfoza. Istota tego zjawiska polega na rozbudowaniu lub redukcji niektórych części ciała w stosunku do innych. Skorelowane to jest ze zmianami temperatury i tym samym lepkości, a ma na celu zwiększenie pływalności i przedłużenie okresu przebywania w odpowiedniej warstwie. Cyklomorfoza zachodzi jedynie u tych gatunków, gdzie w cyklu rocznym występuje po sobie kilka pokoleń. Klasyczny przykład cyklomorfozy opisany został u wioślarki Daphnia cucullata (rys. 27). Zasadniczym zmianom ulega u niej kształt głowy. Osobniki letnie mają głowę wydłużoną hełmiasto, natomiast zimowo-wiosenne są mniejsze z zaokrągloną głową. Tak więc okazy letnie z hełmami są lepiej przystosowane do przeciwstawiania się opadaniu w mniej lepkiej wodzie w tym okresie. Zjawisko cyklomorfozy obserwowane jest u niektórych pierwotniaków, wrotków i skorupiaków oraz, z fitoplanktonu, u bruzdnic.
Rys. 27. Cyklomorfoza u Daphnia cucullata (przy rysunkach podane są daty)
Oprócz producentów i konsumentów w planktonie obecni są reducenci. Ta grupa ekologiczna budowana jest przez bakterie i grzyby wodne. W skład bakterioplanktonu wchodzą przedstawiciele prawie wszystkich rzędów z gromady Bacteria. Pod względem morfologicznym komórki bakterii wodnych są kuliste, cylindryczne lub śrubowate, również spotyka się kształt nitkowaty i stylikowaty. Nitki te mogą być pojedyncze, nierozgałęzione lub rozgałęzione, albo skupione w wiązki. Nienitkowate formy mogą tworzyć skupienia kuliste, owalne, gwiaździste, płytkowate. W jeziorach, szczególnie eutroficznych, dominują nieprzetrwalnikujące pałeczki. W czystych jeziorach ich liczba jest mniejsza na korzyść bakterii przetrwalnikujących. W jeziorach dystroficznych te ostatnie mogą nawet przewyższać liczbę bakterii nieprzetrwalnikujących. W rzekach skład bakterii uzależniony jest od intensywności spływu wody z dorzecza. Przy dużym spływie odsetek bakterii glebowych będzie znaczny. W wielu jeziorach przeważają bakterie gramujemne, również większość bakterii morskich to bakterie gramujemne. Zasadniczo bakterie morskie są halofilami, którym do optymalnego rozwoju potrzebny jest w środowisku chlorek sodu. Obok bakterii heterotroficznych, które w wodach naturalnych przeważają nad innymi typami, notuje się bakterie foto- i chemoautotroficzne. Bakterie fotoautotroficzne występują głównie w dostatecznie naświetlonych rejonach, gdzie obecny jest siarkowodór, zaś chemoautotroficzne także w warstwach głębszych. Stosunki liczebności bakterii w wodzie ściśle korelują z zawartością materii organicznej. Dlatego też w rozkładzie pionowym maksimum bakterii przypada w warstwie podpowierzchniowej, nieco poniżej maksimum biomasy fitoplanktonu. Również w cyklu rocznym zmienność liczebności bakterii heterotroficznych współgra ze szczytami rozwojowymi fitoplanktonu.
W badaniach planktonu podstawowym zadaniem jest wydobycie go z wody poprzez zastosowanie takiego przyrządu, który umożliwiłby odsączenie z wody rozpuszczonej w niej zawiesiny. W 1845 roku Müller użył do tego celu skonstruowanej przez siebie siatki, stosując jako materiał cedzący gazę służącą w przemyśle młynarskim do przesiewania mąki (stąd często mówi się "gaza młynarska"). Potem pojawiło się wiele ulepszonych typów siatek planktonowych, łącznie z modyfikacjami, które dawały materiał do analizy ilościowej. Takimi były siatka planktonowa Hensena, stosowana do połowu planktonu w morzu, oraz siatka Apsteina używana w zbiornikach słodkowodnych (rys. 28). Siatki te jako narzędzia stosowane do połowów ilościowych obarczone są dwiema grupami błędów. Pierwsza to trudności ze ścisłym obliczeniem objętości wody cedzonej przez sieć planktonową. Druga grupa błędów związana jest z trudnościami w pozyskaniu wszystkich organizmów zasiedlających pobieraną próbę wody - drobne formy przechodzą przez oka sieci, natomiast duże, o znacznym stopniu aktywności ruchowej, unikają wlotu siatki planktonowej. Do wyeliminowania tych błędów, a szczególnie pierwszego, służą stosowane od dość dawna czerpacze wody (batometry) (rys. 29). Unowocześnienie konstrukcyjne batometrów szło w kierunku zminimalizowania wirów powstających przy ich opuszczaniu do wody. Uzyskiwane z batometrów próby wody są odsączane metodą filtracji lub wirowania. Aktualnie powszechnie stosowana jest metoda osadzania się zawiesiny planktonowej w specjalnych komorach sedymentacyjnych, zastosowanych po raz pierwszy przez Utermöhla na przełomie lat dwudziestych i trzydziestych. Stosowanie tej metody wymaga używania tzw. mikroskopu odwróconego, pozwalającego na oglądanie od dołu osadzonych na dnie komory sedymentacyjnej organizmów.
Rys. 28. Różne typy siatek planktonowych oraz sploty gazy młynarskiej
Rys. 29. Różne typy czerpaczy wody, tzw. batometrów a - flaszka Meyera, b - czerpacz Ruttnera, c - czerpacz Patalasa 1 - wieczko górne, 2 - zatrzask, 3 - górny kołnierz zewnętrzny, 4 - zawiasy uchwytów, 5 - śruba regulująca siłę zatrzasku, 6 - zawiasy wieczka, 7 - sprężyna zatrzasku, 8 - uchwyt, 9 - szybka, 10 - uchwyt termometru, 11 - gwintowany otwór do wkręcania w niego rurki odpływowej, 12 - wieczko dolne, 13 - dolny kołnierz wewnętrzny
Każda z frakcji wielkościowych planktonu wymaga specyficznych metod poboru prób i ich obróbki oraz specjalistycznego sprzętu do badań.
Nekton
Nekton, w odróżnieniu od planktonu, obejmuje organizmy, które dzięki dobrze rozwiniętym narządom ruchu mogą dowolnie zmieniać miejsce przebywania, przemieszczając się w pionie i w poziomie. Zdolne są także do pływania pod prąd.
W skład nektonu wchodzą ryby i niektóre skorupiaki, a w morzach oprócz tych dwóch grup dochodzą głowonogi, delfiny, kaszaloty, walenie. Wszyscy przedstawiciele tej formacji ekologicznej wykazują specyficzne przystosowania anatomiczne ułatwiające poruszanie się w wodzie poprzez zmniejszanie tarcia. Są to: opływowy kształt ciała, gładka powierzchnia, pokryta śluzem lub tłuszczem.
Ze względu na zajmowany typ siedliska wyróżnia się kilka frakcji nektonu, np. limnonekton - nekton wód stojących, potamonekton - nekton potoków i rzek. W zależności od rejonu zasiedlenia dzieli się, szczególnie nekton morski, na nekton pelagiczny, zasiedlający toń wodną otwartej przestrzeni, oraz nekton nerytyczny związany z płytkimi wodami przybrzeżnymi.
Rozsiedlenie nektonu uzależnione jest przede wszystkim od stratyfikacji termicznej i tlenowej oraz obecności odpowiedniego pokarmu.
Przyjmując za kryterium rejon zasiedlenia oraz rodzaj pokarmu, ryby jeziorne można podzielić na kilka kategorii:
- pelagiczne + planktonożerne (sielawa, stynka, ukleja) i drapieżne (sandacz, troć);
- głębinowo-denne (sieja, leszcz, brzana);
- przybrzeżno-denne + wszystkożerne (karp) i drapieżne (węgorz, okoń);
- przybrzeżne + roślinożerne (płoć, lin, karaś) i drapieżne (szczupak, miętus).
Do ryb w rzekach podział ten nie odnosi się. Tam odpowiednie gatunki grupują się w poszczególnych odcinkach biegu rzeki. I tak w odcinku źródlanym dominują łososiowate z pstrągiem na czele. Odcinek potokowy określa się mianem krainy brzany, a rzeka niżowa to kraina leszcza.
Charakterystyczną cechą biologii ryb są wędrówki związane z rozrodem i żerowaniem. Jest to zjawisko typowe dla ryb morskich. Wiele gatunków ryb morskich wykonuje bardzo odległe wędrówki. Ryby ze względu na stopień przywiązania do środowiska dzieli się na dwie grupy. Są to ryby dwuśrodowiskowe (amfibiotyczne), odbywające odległe wędrówki rozrodczo-żerowiskowe, przebywające w ich trakcie w środowisku morskim i słodkowodnym, oraz ryby jednośrodowiskowe (monobiotyczne), związane całe życie z jednym typem środowiska. Wśród ryb dwuśrodowiskowych wyodrębnia się dwie grupy:
- potamotoca (anadromiczne), ryby rozradzające się w wodach śródlądowych, do nich należy łosoś, żerujący w morzu, a na tarło wędrujący w górę rzeki do jej źródła;
- thalassotoca (katadromiczne), ryby rozradzające się w morzu, tak jak węgorz, który żeruje w rzekach i jeziorach, na tarło wędruje do Morza Sargassowego, tam ginie, a młode węgorzyki w postaci larw unoszonych biernie przemierzają Atlantyk, aby po dwóch latach dotrzeć do brzegów Europy. Ryby jednośrodowiskowe mogą być albo słodkowodne, albo morskie. Te ostatnie dzieli się na osiadłe, praktycznie nie wędrujące, jedynie nieco przemieszczające się (np. flądry), oraz na wykonujące dość znaczne wędrówki sezonowe o charakterze tarliskowo-żerowiskowym (np. śledzie w Morzu Norweskim).
Metody połowu nektonu do badań nie osiągnęły poziomu takiego jak poboru planktonu. Związane jest to z rozmiarami organizmów nektonowych. Spowodowało to przewagę badań populacyjnych nektonu, dla których potrzebne są próby o charakterze statystycznym. Zwykło się je uzyskiwać przy użyciu różnego rodzaju sieci i włoków (rys. 30).
Rys. 30. Przykłady przyrządów służących do połowu nektonu a - sieć pławnica, b - włok, c - okrężnica.
Neuston
Neuston jest formacją drobnych organizmów zasiedlającą warstwę styku wody i atmosfery. Ciśnienie atmosferyczne oraz wynikające z oddziaływania na siebie biegunowo naładowanych cząstek wody, siły spójności dają dość duże napięcie powierzchniowe. Powoduje ono wytworzenie się, dającej pewien opór przy przebijaniu, elastycznej błonki. Przyczepiają się do niej drobne organizmy tworzące zbiorowiska neustonowe. Zbiorowiska tworzą się przy spokojnej, bezwietrznej pogodzie w postaci błoniastego filmu, niekiedy zabarwionego na kolor zielony, żółty, brunatny lub fioletowy.
W skład zbiorowisk neustonu wchodzą bakterie, grzyby (workowce, a przede wszystkim glonowce), z glonów okrzemki, różnowiciowce, zielenice, eugleniny, ponadto różne bezbarwne wiciowce oraz pierwotniaki. Wszystkie one mają przystosowania anatomiczne i fizjologiczne do bytowania w specyficznym środowisku błonki powierzchniowej, gdzie narażone są na niekorzystne promieniowanie i szybkie zmiany temperatury. Zazwyczaj przed tymi niekorzystnymi czynnikami organizmy neustonowe chronią się wytwarzając grubą błonę lub ochronne domki (rys. 31). Zmiana intensywności promieniowania powoduje niekiedy zmianę rozmieszczenia barwników, np. przy silnym naświetlaniu na górnej powierzchni komórki będzie występował barwnik czerwony, natomiast przy słabym - zielony. Czasami zabarwienie osłonki powierzchni pochodzić może od pyłów i sadzy kominowej, jest to wtedy tzw. trypto-neuston.
Rys. 31. Organizmy neustonowe 1 - Chromulina rosanoffi, 2 - Botrydiopsis arhiza, 3 - Nautococcus emersus, 4 - Lampropedia hyalina, 5 - Navicula sp., 6 - Codonosiga botrytis, 7 - Arcella sp.
Ze względu na umiejscowienie neustonu w stosunku do błonki powierzchniowej, wyróżnia się epineuston - organizmy występujące ponad nią i hyponeuston - organizmy przyczepione do błonki od spodu. Ze względu na skład mówi się o fitoneustonie, czyli organizmach roślinnych, i zooneustonie - tworzonym przez organizmy zwierzęce. Na skład neustonu ma wpływ typ zbiornika wodnego - inny będzie w stawach, inny w kałużach, a jeszcze inny w zakolach rzek. W wielu wypadkach, wtedy gdy wiatr zmąci powierzchnię wody, organizmy neustonowe przechodzą okresowo w skład planktonu. Jednak masowo mogą rozwijać się jedynie na powierzchni wody, na granicy fazy wodnej i powietrznej.
Zbieranie organizmów neustonowych jest znacznie prostsze niż łowienie planktonu. Zbiera się je na szkiełka nakrywkowe lub inne podobne. Trzymane w szczypcach szkiełko kładzie się ostrożnie na powierzchni błonki, a następnie nieco skośnym ruchem ostrożnie podnosi się je wraz z przyklejonymi do niego organizmami neustonowymi. Można je również zbierać używając wygiętej nieco ezy mikrobiologicznej. Tak zebraną próbę neustonu utrwala się parami kwasu osmowego lub parami jodu.
Pleuston
Z warstwą powierzchniową wody związana jest większa liczba gatunków niż tylko te, których egzystencja uzależniona jest od zdolności przyczepu do błonki powierzchniowej. One to tworzą formację pleustonu, budowaną przede wszystkim przez nie zakorzenione, pływające pod powierzchnią rośliny. Niektórzy autorzy wliczają do tej formacji również zwierzęta, np. owady poruszające się po powierzchni ruchem ślizgowym, a w morzu rurkopławy powierzchniowe (np. żeglarz portugalski i żagielek). Mimo różnych poglądów na skład gatunkowy tej formacji jej podstawę stanowią rośliny, które częściowo (szczególnie pędy kwiatowe) wystają ponad wodę, zaś system korzeniowy zwisa swobodnie w wodzie. Będąc nie przytwierdzone do podłoża są biernie przesuwane ruchem wody i wiatru. Mogą niekiedy tworzyć wyspy pływające lub zbite darnie, odcinając przez to dopływ promieniowania i hamując rozwój fitoplanktonu w zbiorniku wodnym. U roślin pleustonowych wykształcone są specjalne przystosowania służące do utrzymania się na powierzchni wody. Polegają na zmniejszeniu masy właściwej ciała przede wszystkim poprzez wykształcenie dużych przestworów powietrznych.
Rośliny o cechach wyżej opisanych określa się nazwą pleustonu właściwego. Do nich zalicza się przede wszystkim gatunki rzęs, ponadto żabiściek, rogatek itp. W zbiornikach klimatu tropikalnego różnorodność gatunkowa pleustonu jest znacznie większa. Oprócz wymienionych form wyróżnić można jeszcze pleuston okresowy - meropleuston. Są nim oderwane części roślin zakorzenionych, np. gałązki moczarki, wywłócznika i różnych gatunków rdestnic.
Zbiorowisko pleustonowe najbardziej rozwija się w małych, zacisznych zbiornikach. Szczególnie licznie występuje w płytkich zatokach jeziornych, starorzeczach i dołach potorfowych. Do zbierania roślin pleustonowych stosuje się proste przyrządy, takie jak kotwiczki lub specjalnie przystosowane do tego celu grabie.
Bentos
Formacja ta obejmuje wszystkie organizmy, które w swoim zasadniczym okresie rozwoju, jako formy dorosłe, żyją osadzone na dnie lub innym podłożu, lub są z dnem ściśle związane.
Ze względu na skład wyróżnia się bentos roślinny - fitobentos i zwierzęcy - zoobentos.
W składzie organizmów bentosowych wyróżnia się wiele form życiowych, zróżnicowanych między sobą trybem życia i stopniem związania ze środowiskiem. Biorąc pod uwagę to kryterium wyróżniono cztery kategorie bentosu: rhizobentos, haptobentos, herpobentos i nektobentos.
Rhizobentos obejmuje zakorzenione rośliny, a więc makrofity. Rejon jego występowania to litoral, gdzie docierające ilości światła pozwalają na przebieg fotosyntezy. Rhizobentos klasycznie wykształcony jest w zbiornikach jeziornych, zaś w rzekach zależy od intensywności przepływu wody. W morzu roślin naczyniowych jest niewiele.
Rośliny naczyniowe tworzące zasadniczy składnik rhizobentosu wykazują specyficzne przystosowania do środowiska. Dotyczą one przede wszystkim sposobu przytwierdzania się do podłoża - w tym celu wytwarzają rozległe systemy kłączy, rozłogów i korzeni, co uniemożliwia ich łatwe wyrwanie. Rośliny podwodne, w związku z ruchem mas wodnych, muszą wykazywać elastyczność, dlatego mają wiotkie pędy. Cecha ta wynika ze słabego wykształcenia tkanki mechanicznej, ułożonej centralnie. Wiele z nich tworzy długie, taśmowate liście. Następną ważną cechą przystosowawczą jest wytwarzanie w łodygach i liściach dużych przestworów międzykomórkowych, w których gromadzi się powietrze. Zmagazynowane tam gazy są wykorzystywane w procesach życiowych, w fotosyntezie i oddychaniu. Ponadto nadają liściom lekkość i pływalność. Dzięki tym przestworom liście pływające grążeli i innych roślin nie toną.
Wśród makrofitów zakorzenionych wyróżnia się cztery kategorie form życiowych:
- amfifity - rośliny ziemnowodne, bytujące w strefie przejściowej między lądem a wodą; zalicza się do nich wiele traw, turzyc, jaskrów itp.;
- helofity - rośliny bagienne, zakorzenione w podłożu stale zalanym, natomiast części zielone i kwiaty są wynurzone; do nich zalicza się m.in. trzcinę, pałkę, sit, tatarak;
- nimfeidy - rośliny o liściach pływających na powierzchni wody, do grupy tej należy przede wszystkim lilia wodna i grążel;
- elodeidy - rośliny całe zanurzone pod powierzchnią wody, takie jak moczarka kanadyjska, wywłócznik i kilka gatunków rdestnic.
Wymienione cztery grupy form życiowych makrofitów wodnych układają się najczęściej strefowo. Na samym brzegu znajduje się pas amfifitów, dalej w stronę wody występują helofity, następną strefę tworzą nimfeidy, zaś najgłębiej rosną elodeidy, które w płytkich zbiornikach mogą zajmować całą powierzchnię dna. Wśród elodeidów, w niektórych typach zbiorników wodnych ważną rolę odgrywają ramienice, a z mchów Fontinalis.
Flora makrofitów wodnych, podobnie jak lądowych, badana jest metodami fitosocjologicznymi. W zależności od charakteru podłoża i chemizmu wody wyróżniono kilkanaście zespołów, z wieloma wariantami i facjami. W limnologii funkcjonują uproszczone systemy klasyfikacji makrofitów. Służą one do opisu różnych typów jezior z uwzględnieniem poszczególnych gatunków makrofitów jako wskaźników. Dla jezior polskich wykaz wskaźników roślinnych zaproponował Bernatowicz (tab. 6).
Tabela 6.
Wykaz limnologicznych wskaźników roślinnych
Źródło: Mikulski J.S. Biologia wód śródlądowych. Warszawa, PWN 1974.
W limnologii rybackiej roślinność wodną dzieli się na dwa typy: twardą i miękką. Roślinność twarda to przede wszystkim helofity, bujnie zarastające zbiorniki i wiążące znaczne ilości biogenów. Ich rozkład zachodzi dość wolno, stąd też powrót do ekosystemu biogenów zmagazynowanych przez nie jest utrudniony. Roślinność miękka, czyli zanurzona, z punktu widzenia rybackiego jest elementem korzystnym - jest miejscem schronienia dla wielu drobnych zwierząt, które na niej żerują, stając się jednocześnie pożywieniem dla ryb. Ponadto są dogodnym miejscem rozrodu zwierząt, w tym również miejscem tarliskowym dla ryb.
Haptobentos to organizmy nie związane z dnem, wymagające jednak stałego podłoża. Stanowić je może element dna, np. pale, umocnienia, duże głazy, wraki łodzi lub statków, albo też element żywy jak rośliny, czy zwierzęta denne. Frakcję tę tworzą rośliny lub zwierzęta, które przylegają bądź przyczepiają się do podłoża lecz nigdy nie wchodzą weń systemem korzeniowym, tak jak makrofity wodne. Do haptobentosu zalicza się wiciowce roślinne, osiadłe okrzemki, a przede wszystkim glony nitkowate, zaś ze zwierząt głównie gąbki i mszywioły oraz osiadłe pierwotniaki i wrotki. Te drobne organizmy rosnące na wolnej powierzchni obiektów zanurzonych w wodzie określane są przez wielu autorów nazwą peryfitonu. Jest więc to zbiorowisko o charakterze mikrocenozy, z wyraźnie zarysowanymi powiązaniami troficznymi. Podstawą peryfitonu są producenci czyli glony. Ponadto liczne są tam bakterie. Natomiast konsumentami są tacy stali mieszkańcy peryfitonu jak korzenionóżki, osiadłe i pływające wrotki, nicienie, brzuchorzęski, skąposzczety, także wioślarki i larwy ochotek, a nawet młode larwy jętek. W morskiej faunie osiadłej spotyka się przedstawicieli koralowców, pierścienic, czułkowców i osłonic. Również osiadły tryb życia, spośród skorupiaków, prowadzą wąsonogi, np. pąkle i kaczenice. Do tych morskich porośli zalicza się także niektóre małże, np. omułka i ostrygę.
Biocenozy peryfitonu niekiedy wykazują specyficzną strukturę - zbudowane są jakby piętrowo. Pierwszą warstwę tworzą glony nitkowate chwytnikami przytwierdzone do podłoża. Drugą również glony nitkowate, ale bez chwytników - oplatają one poprzednie. Trzecie piętro to okrzemki, które osadzają się na poprzednich, a czwartym elementem strukturalnym są zwierzęta i glony nie przytwierdzone, przemieszczające się między poprzednimi piętrami. Skład gatunkowy zbiorowisk peryfitonowych zależy od chemizmu wody, a nie jest związany z gatunkiem rośliny, czy zwierzęcia. Charakter mechaniczny podłoża może jednak warunkować wytworzenie się peryfitonu. Na powierzchni gładkiej pokrytej śluzem tworzy się on trudniej niż na powierzchni chropowatej.
Herpobentos, określany nazwą bentosu właściwego (eubentos), obejmuje organizmy zwierzęce, żyjące na dnie lub w dnie, mogące się po nim lub w nim przemieszczać. Tworzą tę formację gatunki należące do wielu grup taksonomicznych, poczynając od pierowtniaków do najbardziej uorganizowanych bezkręgowców włącznie. Pod względem wielkości są to bardzo różne formy, stąd też wyróżnia się trzy frakcje wielkościowe: makrobentos, o rozmiarach powyżej 2 mm, mezobentos (lub meiofauna), w zakresie wielkości 0,2 (0,1)-2mm, mikrobentos poniżej 0,1 (0,2) mm. Dolna granica wymiarów meiofauny jest niejednolita i różna według różnych autorów, od 0,1 do 0,5 mm. Organizmy tworzące herpobentos, w związku z ich trybem życia, wykazują specyficzne przystosowania pozwalające na przebywanie na dnie.
Organizmy bentosowe, w celu uniknięcia wypychania ich ku powierzchni wody, są zaopatrzone w ciężkie szkielety wapienne; chityna jako typowy budulec szkieletu wielu organizmów planktonowych jest u nich zwykle przesycona wapniem. Wapienne szkielety występują u licznych małży, ślimaków, szkarłupni i skorupiaków, szczególnie wtedy, gdy są wystawione na niszczące działanie fal. Wiele form ma nawet możliwość wnikania w dno, co daje im daleko idące zabezpieczenie.
Ze względu na stopień związania się z podłożem wyróżnia się kilka form życiowych:
- Formy zagrzebujące się. Zalicza się tu liczne gatunki z wielu grup taksonomicznych, przede wszystkim spośród małży, krabów i robaków.
- Formy budujące domki. Do fauny bytującej na osadach miękkich należy sporo gatunków wytwarzających z mułu i wydzielanego przez siebie śluzu domki w formie rurek. W wodach słodkich typowym przykładem są rureczniki (Tubifex). W morzach takie domki tworzą liczne wieloszczety, obunogi oraz wiele innych mieszkańców strefy przybrzeżnej.
- Formy drążące. Wiele gatunków osiadłych ma zdolność przenikania w twarde podłoże. Jedne z nich, np. niektóre małże (skałotocze), używają ostrego brzegu skorupy jako swoistego świdra, względnie trawią podłoże, zwykle wapienne, za pomocą wydzielanych przez siebie kwasów. Inne, np. świdrak okrętowy, znany niszczyciel drewnianych okrętów, budowli i umocnień portowych, ma muszlę skręconą na kształt prawdziwego świdra.
- Formy denne ruchome (wagilne). Formy te trybem życia są ściśle związane z dnem, jednak mogą się swobodnie poruszać po jego powierzchni. Bentos wagilny tworzą przedstawiciele licznych grup zwierzęcych: jamochłonów, robaków, pierścienic, stawonogów, mięczaków, szkarłupni itp.
Ze względu na charakter mechaniczny dna wyróżnia się trzy podstawowe typy biocenoz bentosowych:
- Liton: wykształca się na dnie kamienistym i składa się ze zwierząt mających zdolność silnego przywarcia do podłoża. Zwierzęta te przylegają do podłoża całą powierzchnią ciała albo zaopatrzone są w przyssawki lub mają silne kończyny zakończone pazurami.
- Psammon: biocenoza dna piaszczystego. Zwierzęta tej biocenozy muszą być przystosowane do sypkości podłoża; duże posiadają pancerze chroniące je przed zgnieceniem przy przemieszczaniu się piasku, a małe muszą być tak małe, aby mogły żyć w przestrzeniach między ziarnami piasku. Forma dna piaszczystego nie jest bogata w gatunki.
- Pelon: obejmuje zbiorowiska zwierząt żyjących na dnie mulistym, zbudowanym z osadów mineralnych, drobnocząsteczkowych lub organicznych. Różne rodzaje dna mają bardzo obfitą i zróżnicowaną faunę denną. W warstwie przybrzeżnej różnorodność gatunkowa jest znacznie większa niż w rejonach głębszych.
Nektobentos obejmuje organizmy przebywające w strefie ponad dnem. Są one z dnem związane, mogą nawet przebywać na nim przysypane piaskiem, jednakże cechą specyficzną dla nich jest możliwość swobodnego pływania, co prawda tylko tuż ponad dnem, ale dzięki temu mogą się łatwiej przemieszczać i zmieniać miejsce pobytu. Do tej formacji należą larwy wodzieni (Chaoborus) oraz ryby płaskie, płazy itp.
Metody poboru prób bentosu są różnorodne. Zależą od celu badań, a więc od tego czy mają one służyć analizie jakościowej, czy ilościowej. Również ze względu na charakter dna skonstruowano wiele specjalistycznych przyrządów służących do zbioru organizmów bentosowych (rys. 33). Podstawowym narzędziem, służącym do zorientowania się w składzie bentosu jest draga. Jest ona rękawem z tkaniny łatwo przepuszczającej wodę, ciągnionym na lince po dnie. Przyrządy, za pomocą których zbiera się materiał służący do oceny liczebności, to różnego rodzaju czerpacze dna (typy: Ekman do badań jeziornych; Petersen do badań morskich). Do zbioru drobnego zoobentosu (meiofauny i mikrofauny) na dnie piaszczysto-mulistym i mulistym służą czerpacze rurowe. Ponadto stosuje się metody fotograficzne i telewizyjne, zaś niezawodne badania bentosu można prowadzić metodą swobodnego nurkowania lub obserwacji z batyskafu.
Rys. 33. Przyrządy do zbioru bentosu a - dragi, b - czerpacz dna Ekmana: 1 - otwarty, 2 - zamknięty, c - czerpacz dna Petersena: 1 - otwarty, 2 - zamknięty, 3 - opuszczany z burty statku
Zbiorniki wodne
Morza i oceany
W środowisku morskim wydziela się dwa zasadnicze obszary, środowiskowo bardzo zróżnicowane. Jest to obszar toni wodnej, zwany dziedziną pelagiczną lub pelagialem oraz obszar dna morskiego, zwany dziedziną bentoniczną lub bentalem (rys. 33).
W pelagialu wyróżnia się dwa rejony: nerytyczny i oceaniczny.
Rejon nerytyczny obejmuje pas wód przybrzeżnych aż do linii, gdzie głębokość nie przekracza 200 m. Dno tego rejonu określane jest mianem szelfu kontynentalnego.
Rejon oceaniczny obejmuje wody otwarte poza rejonem nerytycznym - od powierzchni aż do największych głębokości oceanicznych. W rejonie tym wydziela się następujące warstwy: epipelagial - od powierzchni do głębokości 200 m, mezopelagial - w zakresie głębokości 200-1000 m, batypelagial - od 1000 do 4000 m, abysopelagial - poniżej 4000 m.
W górnej części epipelagialu wykształcona jest warstwa eufotyczna. Zasięg jej jest różny w różnych rejonach, jednakże nawet w bardzo czystych wodach tropikalnych, gdzie ilość zawiesiny jest nieduża, rzadko sięga ona poniżej głębokości 100 m (możliwe maksimum: 150 m w Morzu Sargassowym). Miąższość mezopelagialu odpowiada mniej więcej zasięgowi warstwy dysfotycznej. Mezopelagial zamieszkały jest przez gatunki ryb o niezwykle dużych i czułych oczach, zdolnych do wykrywania światła 100-krotnie słabszego niż to, które może wykryć oko ludzkie. Innymi ważnymi mieszkańcami tej warstwy są organizmy wykazujące zdolność bioluminescencji. Są to zwłaszcza krewetki, kalmary i ryby. Prawie 80% gatunków zwierząt żyjących w mezopelagialu posiada narządy świetlne, tzw. fotofory, w których produkowane jest "zimne światło". Batypelagial i abysopelagial obejmuje warstwa afotyczna, w której żyje wiele ryb zupełnie ślepych, ponieważ wobec braku światła oczy stają się zbyteczne. Gatunki żyjące w warstwie afotycznej są drapieżnikami, charakteryzują się wydłużonym ciałem oraz dużą głową z nieproporcjonalnie w stosunku do rozmiarów ciała wykształconą paszczą. Mają bardzo dobrze rozwinięte urządzenia ostrzegawcze i specjalne organy do zdobywania pożywienia, które jest w tej strefie niezmiernie rozproszone.
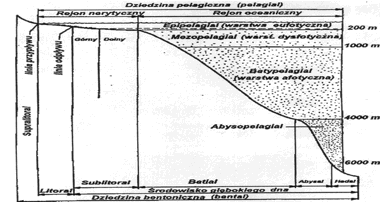
Rys. 33. Podział środowiska morskiego
Dziedzinę bentoniczną dzieli się na dwa rejony, w których na podstawie różnych kryteriów wyodrębnia się kilka dalszych jednostek. Tymi podstawowymi rejonami są: litoral i środowisko głębokiego dna.
Litoral jest to strefa obejmująca obszar zasięgu pływów, zaś środowisko głębokiego dna obejmuje pozostały obszar dna oceanicznego. Ponad litoralem wyodrębnia się strefę supralitoralu, pospolicie określaną jako strefa oprysku, czyli rejon brzegu okresowo zalewany, np. podczas silnych sztormów. Poniżej litoralu, czyli poniżej poziomu odpływu aż do głębokości 200 m wyodrębnia się strefę sublitoralu, który dzielony jest na górny i dolny. Górny sublitoral sięga aż do granicy zasięgu występowania roślin, co uzależnione jest od ilości światła docierającego, pozwalającego na proces fotosyntezy. Na ogół jest to głębokość do około 50 m, ale w rejonach o dużym zmętnieniu jest to poziom znacznie płytszy. Litoral i sublitoral są wykształcone na szelfie kontynentalnym, czyli półce będącej przedłużeniem lądu w kierunku morza. Dalszą strefą jest batial, sięgający do głębokości 4000 m, odpowiada on tzw. stokowi kontynentalnemu, czyli stromej ścianie, na której kończy się płyta kontynentu. Poniżej do głębokości 6000 m rozciąga się obszerna, przeważnie płaska, strefa abysalna, obejmująca ponad 80% dna oceanicznego. W niektórych rejonach głębiej wykształca się specyficzne środowisko rowów oceanicznych określane nazwą hadalu lub czasami ultraabysalu.
Znajomość zasiedlenia wszechoceanu przez organizmy jest słaba, ze względu na jego olbrzymią przestrzeń. Wiadomo jednak, że na ponad milion gatunków zwierząt w faunie światowej jedynie 16% żyje w oceanach, z tego przytłaczająca większość, bo około 98% zasiedla dno oceaniczne, a pozostałe 2% występuje w otwartych wodach oceanów. Oceany zasiedlone są nierównomiernie. Spowodowane jest to specyfiką wymagań ekologicznych poszczególnych gatunków, jak i różnorodnością i stopniem wykształcenia czynników środowiskowych warunkujących rozwój życia. Te ostatnie zależą zasadniczo od szerokości geograficznej oraz głębokości.
Fitoplankton oceaniczny budują okrzemki (Bacillariophyeae), bruzdnice (Pyrrophyta), wiciowce wapienne (Coccolithineae) i krzemionkowce (Silicolagallatae). Zaznacza się zróżnicowanie przestrzenne w rozmieszczeniu fitoplanktonu. Okrzemki są typowe dla wód zimnych, zaś bruzdnice i wiciowce dla ciepłych. Zooplankton obejmuje gatunki pierowtniaków (Protozoa), jamochłonów (Coelenterata), żebropławów (Ctenophora), pierścienic (Annelida), strzałek (Chaetognatha), mięczaków (Mollusca), stawonogów (Arthropoda) i szkarłupni (Echinodermata). Te ostatnie reprezentowane są w planktonie przez formy larwalne. Prócz tego w planktonie występują pierwotniejsze formy strunowców - osłonice (Tunicata) oraz ikra i stadia młodociane ryb. Zwierzęta planktonowe występują prawie we wszystkich strefach pelagialu. Czynnikiem limitującym dla nich nie jest światło, jak w przypadku fitoplanktonu, a zasobność wody w pokarm oraz temperatura. Do nektonu morskiego zalicza się ryby, kalmary, morskie gady i ssaki.
Fitobentos morski obejmuje glony z trzech gromad - zielenic (Chlorophyta), brunatnic (Phaeophyta) i krasnorostów (Rhodophyta) oraz nieliczne gatunki roślin naczyniowych. Występują one we wszystkich morzach, jednak ich rozmieszczenie uzależnione jest od strefy klimatycznej, głębokości wody i innych czynników ekologicznych. W skład zoobentosu wchodzą gatunki pierwotniaków (Protozoa), gąbek (Poriferia), koralowców (Anthozoa), ukwiałów (Actinaria) wstężniaków (Nemertini), pierścienic (Annelida), mięczaków (Molusca), stawonogów (Arthropoda), jeżowców (Echinoidea), liliowców (Crinoidea), rozgwiazd (Asteroidea) i wężowideł (Ophiuroidea). Największa różnorodność zoobentosu występuje w strefie litoralu. Im głębiej, tym liczba gatunków maleje. Wiele z nich ma szeroki zasięg występowania, a te, które występują w rejonach głębokiego dna, wykazują dużą tolerancję na zmiany ciśnienia.
Jeziora
Ogólna charakterystyka jezior
Jezioro jest to zbiornik wody wykazujący pionową strefowość czynników ekologicznych, wykształcony w naturalnym zagłębieniu na powierzchni Ziemi, nie mający bezpośredniego, szerokiego połączenia z morzem.
Jeziora dzieli się na bezodpływowe, czyli zamknięte, oraz odpływowe, czyli otwarte. Jeziora bezodpływowe, szczególnie w strefie klimatu ciepłego, gdzie parowanie jest znaczne, wykazują podwyższoną zawartość elektrolitów. W konsekwencji jeziora takie mają wody słone, niejednokrotnie o zasoleniu wyższym niż oceaniczne. Efektem tego są specyficzne stosunki biocenotyczne, z właściwą im fauną i florą. Jeziora otwarte, czyli odpływowe (z jeziora wypływa rzeka), lub przepływowe (do jeziora wpływa i wypływa rzeka) są typowe dla klimatu umiarkowanego i wilgotnego (np. jeziora bałtyckie).
Jeziora ze względu na ich pochodzenie dzieli się na kilka kategorii. Jeziora tektoniczne, powstałe na skutek pęknięć w skorupie ziemskiej lub przesunięć masywów krystalicznych, są najstarsze, zwykle trzeciorzędowe. Typowym przykładem jest tu Bajkał ze swoistą, endemiczną fauną. Dużą grupę stanowią jeziora polodowcowe powstałe w związku z działalnością erozyjną lodowców. W tej grupie wyróżnia się: jeziora moreny dennej, mieszczące się we wklęsłościach pierwotnej równiny denudacyjnej; jeziora moreny czołowej, zamykającej koniec zagłębienia lodowca; jeziora rynnowe, tworzące się w bruzdach drążonych niegdyś przez wody topniejących lodowców; jeziora cyrkowe, zwykle wysokogórskie, wyżłobione przez lodowiec w skale, np. Czarny Staw w Tarach. Kolejnymi kategoriami jezior są jeziora wulkaniczne powstałe w kraterach wygasłych wulkanów i jeziora krasowe wykształcone w zapadliskach na obszarach wapiennych lub gipsowych. Jeziora rzeczne powstały wskutek zamulenia odpływu wód rzecznych, i rozlania się ich poza korytem, lub zatamowania biegu rzeki przez zwały tektoniczne lub zsuwy materiału górskiego. Jeziora przybrzeżne (np. Łebsko, Sarbsko, Gardno) oddzielone są od morza mierzeją wydmową.
Każde jezioro wykształcone jest w zagłębieniu terenu, które nosi nazwę misy jeziornej. W misie jeziornej wyodrębnia się otwartą przestrzeń, czyli toń jeziorną, oraz stoki i dno misy, zaś tuż przy brzegu wykształca się o różnej szerokości platforma lub ławica przybrzeżna (rys. 34). W toni wodnej wydziela się pas wody przybrzeżnej, zalegający nad ławicą przybrzeżną; jest to strefa nerytyczna. Całe śródjezierze określa się nazwą strefy pelagicznej. Na dnie misy jeziornej wydziela się idąc od brzegu: epilitoral - pas ponad zasięgiem zalewania; supralitoral - strefa okresowo zalewana; litoral - strefa dna platformy brzeżnej do granicy występowania makrofitów; sublitoral - strefa wykształcona na stoku platformy brzeżnej, która kończy się na dolnej granicy metalimnionu; profundal - rozległy obszar dna misy jeziornej.
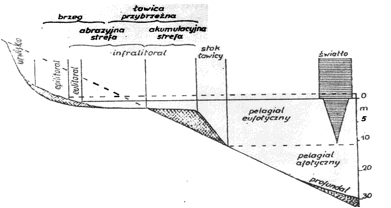
Rys. 34. Podział środowiska jeziornego
Toń wodna, a przede wszystkim strefa pelagiczna, jest tą dziedziną ekologiczną (obszarem), gdzie zachodzą wymienione uprzednio podstawowe procesy hydrobiologiczne. Dotyczy to przede wszystkim specyfiki rozkładu pionowego wszystkich czynników środowiskowych oraz ich zmienności w cyklach dobowych i sezonowych. Efektem tego jest uwarstwienie produktywności biologicznej jeziora i w konsekwencji przewaga procesów katabolicznych (rozkładu) nad anabolicznymi (produkcji) w warstwach głębszych. Pozostaje to w ścisłym związku z charakterem procesów życiowych na dnie jeziora. Strefa pelagiczna odznacza się większą stabilnością procesów i zjawisk ekologicznych niż strefa nerytyczna. W strefie nerytycznej widoczny jest wpływ brzegu i dna - prostym przykładem jest obecność w planktonie form bentosowych. Spowodowane jest to zachodzącym tu intensywnym mieszaniem wody.
Litoral jest najbardziej zróżnicowaną strefą w jeziorze. Wynika to przede wszystkim z wyraźnie zaznaczonej rytmiczności dobowej procesów środowiskowych oraz z dużej różnorodności struktury i składu chemicznego dna. W litoralu jeziornym występują przedstawiciele wszystkich grup roślin i zwierząt wodnych, w przeciwieństwie do pozostałych stref, gdzie wielu grup brak lub reprezentowane są przez nieliczne, wyspecjalizowane gatunki. W litoralu, prócz roślin naczyniowych, licznie występują okrzemki, sinice, zielenice, z bogato reprezentowaną grupą desmidii, oraz bruzdnice i wiciowce. Tworzą one w większości typową dla litoralu formację peryfitonu. Również wśród roślin naczyniowych liczne są drobne zwierzęta, takie jak wioślarki, widłonogi, wrotki, małżoraczki i wodopójki. Litoral jest podstawowym siedliskiem różnych gatunków stułbi, wypławków, nicieni i pierścienic, a także idealnym miejscem pobytu dla ślimaków, zwłaszcza płucodysznych. Ważnym składnikiem fauny litoralu są zwierzęta ziemnowodne, a w tym przede wszystkim, reprezntowane przez wiele grup, owady. Jedne z nich są typowym składnikiem bentosu, np. larwy chruścików lub ochotek, inne zaś swobodnie poruszają się w toni wodnej, bytują na dnie lub powierzchni wody, albo nawet okresowo mogą opuszczać zbiornik. Wśród tych ostatnich licznie reprezentowane są jętki oraz mniej liczne widelnice, larwy ważek, pluskwiaki wodne, drobne chrząszcze, kałużnicowate, zabarwice i inne.
Sublitoral zaczyna się na granicy oddziaływania fal, - pokrywa się to z zasięgiem łąk podwodnych. Sublitoral jest strefą przejściowa między płyciznami litoralu a głębią profundalu. Ograniczony dopływ światła uniemożliwia tu rozwój roślin. Inne czynniki środowiskowe wraz z wzrastającą głębokością wykazują coraz większą stabilność. Ze względu na dobre warunki pokarmowe fauna denna jest w sublitoralu jeszcze ilościowo obfita, ale wraz ze wzrostem głębokości maleje liczba gatunków. Obecne są przede wszystkim mięczaki, pierścienice (skąposzczety i pijawki) oraz larwy owadów. Spośród ślimaków będą tu już skrzelodyszne. Niekiedy w znacznych ilościach obecne są małżoraczki. W sublitoralu mieści się granica między warstwą trofogeniczną a trofolityczną, dlatego też w głębszych jego rejonach odbywają się intensywne procesy rozpadu - destrukcji. Stąd też bogata jest w nim mikrofauna i flora bakteryjna. W wyniku sukcesywnego gromadzenia się opadającej materii organicznej, rosnącej głębokości towarzyszy wzrastająca ilość osadów dennych.
Profundal jest to strefa denna, wyścielona osadami o pochodzeniu głównie organicznym i autochtonicznym. Zalegający nad nim hypolimnion stwarza warunki stabilności czynników środowiskowych. Jest to strefa uboga w tlen, w której zachodzą przede wszystkim procesy rozkładu. Głównymi komponentami fauny profundalnej są skąposzczety (Oligochaeta) i larwy ochotkowych (Chironomidae). Cechą charakterystyczną rozmieszczenia fauny w profundalu jest to, iż występuje ona w skupiskach, między którymi rozciągają się znaczne obszary "pustynne".
Omawiane trzy strefy dna nie są oddzielone ostro od siebie i organizmy bentosowe mogą wędrować z jednej do drugiej. Wędrówki te współgrają z rytmiką zmian sezonowych w środowisku.
Typologia jezior
Jezioro można scharakteryzować poprzez opis cech środowiskowych, wykonując odpowiednie analizy fizyczno-chemiczne wody i poprzez opis zbiorowisk organizmów żywych. Współczesne podziały jezior oparte są na tych czynnikach środowiskowych, które regulują ogólną produkcję biologiczną wód naturalnych. Szwedzki limnolog Naumann wyodrębnił 5 zasadniczych czynników środowiskowych wpływających na wielkość produkcji. Są to: zawartość biogenów, wysycenie gazami, ilość materii organicznej, temperatura i natężenie światła. Ich oddziaływanie na środowisko jest wzajemnie powiązane, jednak stopień wykształcenia w środowisku może być zróżnicowany. Gdy któryś z czynników występuje w dużej ilości to mówi się, że jest w politypie, gdy w małej - w oligotypie, a pośrednio - w mezotypie. Określenie liczbowe poszczególnych typów jest trudne i jest swoiste dla danego rejonu geograficznego.
Przyjęta za Thienemannem klasyfikacja jezior wyodrębnia dwie zasadnicze grupy:
- Jeziora harmonijnie wykształcone, które charakteryzują się równomierną obecnością wszystkich czynników ekologicznych. Czynniki te mogą występować w dużych ilościach, wówczas będą to jeziora eutroficzne, lub w małych ilościach - jeziora oligotroficzne. Stany pośrednie obejmują jeziora mezotroficzne.
- Jeziora jednostronnie wykształcone, gdy jeden z czynników ekologicznych jest w nadmiarze. W grupie tej wyróżnia się kilka typów jezior, w zależności od czynnika, który wywiera zasadnicze piętno, czyli występuje w nadmiarze. Mogą to być między innymi duże ilości: związków humusowych - jeziora dystroficzne, wapnia - jeziora alkalitroficzne, żelaza - jeziora siderotroficzne, glinu - jeziora argillotroficzne, niski odczyn wody (pH) - jeziora acidotroficzne.
Każdy z tych typów jezior ma swoiste stosunki biologiczne, charakteryzujące się specyfiką składu gatunkowego, stosunkami dominacji i liczebnością poszczególnych grup fauny i flory, tak w planktonie, jak i w bentosie. Praktyczny opis cech wyróżniających trzy zasadnicze typy jezior europejskich zawiera tab. 7. Prócz takiego, można powiedzieć środowiskowego podejścia w typologii jezior, zarysował się nurt, w którym elementy biocenozy są podstawą wyróżniania typów jezior. Przykładem takiego podejścia są wskaźniki fitoplanktonowe Thunmarka i Nygaarda oraz inne próby, w których organizmami wskaźnikowymi były poszczególne gatunki zooplanktonu lub fauny dennej.
Tabela 7. Charakterystyka głównych typów jezior
Źródło: Lityński A. Hydrobiologia ogólna. Warszawa, PWN 1952.
Jezioro jako ekosystem dynamiczny zmienia się w czasie. Zmiany te są jednokierunkowe i polegają zwykle na stopniowym zarastaniu. Jest więc to ewolucja związana ze zjawiskiem wzrostu produktywności. Nasilenie i kierunek uzależnione są przede wszystkim od obszaru i charakteru zlewni oraz tego co się w tej zlewni dzieje.
Ewolucja harmoniczna jezior ma postać prostą: jeziora oligotroficzne -> jeziora a-mezotroficzne -> jeziora b-mezotroficzne -> jeziora eutroficzne -> jeziora stawowe -> torfowiska niskie. Istnieje jednak wiele czynników środowiskowych, które ten prosty ciąg w rozwoju jezior modyfikują, np. dopływ humusu może spowodować dystrofię każdego z powyższych typów i w efekcie zarośnięcie jeziora dystroficznego, prowadzące do powstania torfowiska wysokiego.
Zbiorniki zaporowe
Zbiorniki zaporowe to najczęściej sztuczne zbiorniki wodne powstałe przez zagrodzenie doliny rzeki tamą, w wyniku czego następuje spiętrzenie wody. Zbiorniki takie mają swoiste cechy, które różnią je od jezior i rzek. Cechami tymi są:
- asymetria dna - najgłębsze jest przy tamie;
- zmienny poziom wody;
- górna warstwa wody płynie, głębsze warstwy stagnują (zależne od stopnia otwarcia tamy).
Asymetria misy zbiornika zaporowego powoduje nierównomierny rozkład czynników fizycznych i chemicznych oraz biologicznych w zbiorniku. W górnej warstwie panują podobne warunki jak w rzece o szybkim nurcie, a więc równomierny rozkład temperatury, gazów oraz soli mineralnych. W niższych partiach zbiornika wykształca się stratyfikacja taka jak w jeziorach, z warstwą epilimnionu, metalimnionu i hypolimnionu oraz konsekwencjami termicznymi i chemicznymi, które wynikają ze stagnacji letniej.
W okresie początkowym rozwoju zbiornika zaporowego zachodzą specyficzne procesy przemian w glebie i roślinności nagle zalanej wodą. Darń roślinna ulega rozkładowi i mineralizacji. Efektem tego są niekorzystne zjawiska, takie jak zanik tlenu w strefie przydennej i pojawienie się siarkowodoru, zwiększenie się ilości bakterii, podniesienie zawartości żelaza, a nawet manganu. Po jakimś czasie zjawiska te ustają. Ciągle natomiast zachodzić będzie akumulacja zawiesiny nanoszonej wodami wpływającej rzeki. W wyniku tego procesu tworzą się allochtoniczne pokłady osadów dennych, które w zależności od chemicznego charakteru zlewni mogą w wielu przypadkach decydować o troficzności zbiornika.
W zbiorniku zaporowym biocenozy, podobnie jak środowisko, są swoiste. Fitoplankton rozwija się dobrze ze względu na lepsze niż w rzece warunki świetlne, a dzięki temu zooplankton ma doskonałe zasoby pokarmowe. Również bakterie rozwijają się obficie, szczególnie w pierwszym okresie, ze względu na intensyfikację procesów rozkładu materii organicznej. W górnej warstwie zbiornika zdarzają się niekiedy długotrwałe zakwity okrzemek, sinic i zielenic, zaś w zooplanktonie ilość wrotków bywa równa ilości skorupiaków. Głębiej zooplankton składa się przede wszystkim ze skorupiaków. Glony nitkowate, szczególnie zielenice oraz roślinność naczyniowa rozwijają się intensywnie zaraz po zalaniu zbiornika. Rozwój fauny dennej uzależniony jest od procesów kształtowania się charakteru dna. Największym zmianom podlegają biocenozy dna piaszczystego (psammon), gliniastego (argilon) i kamienistego (liton). Intensywnie rozwijają się biocenozy dna mulistego (pelon) oraz peryfiton. Niestabilność tych pierwszych biocenoz wynika ze zmiennego poziomu wody w zbiornikach zaporowych. W ichtiofaunie dla małych zbiorników zaporowych typowe są okoń i płoć, w większych ich miejsce zajmuje leszcz.
Stawy
Brak strefy głębinowej to cecha odróżniająca stawy od jezior. Cecha ta oznacza nieograniczony dostęp światła do dna pozwalający na rozwój roślinności zakorzenionej na całej jego powierzchni. Warunki środowiskowe w stawach są podobne do tych, jakie panują w strefie przybrzeżnej jezior, czyli rejonie nerytycznym. Wahania temperatury są znaczne, a amplituda między dniem i nocą sięga kilku stopni Celsjusza. W stawach nie wykształcają się długotrwałe uwarstwienia termiczne. W lecie przy dnie woda jest chłodniejsza niż na powierzchni, jednak temperatura jest zawsze wyższa od 4oC. W zimie wody przydenne oziębiają się poniżej 4oC.
W związku z dobrymi warunkami do fotosyntezy, wolny dwutlenek węgla zostaje w czasie dnia całkowicie pochłonięty przez rośliny. Powoduje to wytrącanie się z wody szarego węglanu wapnia i osadzanie na roślinach wodnych - rośnie w tym czasie odczyn wody. W cyklu rocznym waha się w stawach zawartość elektrolitów. Wiosną wody roztopowe powodują rozcieńczenie stężenia soli biogenicznych w stawie; ma on więc w tym czasie charakter oligotroficzny. W lecie i jesienią wskutek parowania wody stawowej i dopływu do stawu wód żyznych z pól zwiększa się ilość biogenów i staw przybiera charakter eutroficzny. Stąd też charakterystycznym zjawiskiem letnim w stawach są zakwity glonów, a szczególnie sinic. Wskutek obumierania glonów pochodzących z tych zakwitów i ich osadzania się na dnie, tworzy się gnijąca warstwa mułu, zużywająca znaczne ilości tlenu, co prowadzi najczęściej do całkowitego zaniku tlenu. W stawach nie jest to jednak zjawisko niebezpieczne, gdyż nawet przy słabym wietrze następuje wymieszanie wody do dna i tym samym nasycenie warstw przydennych tlenem. Tak więc warunki w stawach są bardzo zmienne, co praktycznie sprzyja występowaniu organizmów mało wrażliwych na wpływ czynników zewnętrznych (gatunki eurytopowe).
Charakterystyczną formacją dla stawów są rośliny naczyniowe. Rosną one niekiedy zwartymi łanami (np.: turzyce, skrzypy, sitowie, trzcina, pałka wodna), przy czym na podłożu alkalicznym dominuje trzcina; na kwaśnym: turzyce, manna mielec, sity, wełnianki; na podłożu obojętnym, gliniastym - jeżogłówka. W niedużych stawach silnie rozwija się moczarka kanadyjska, jaskry wodne lub rzęsy. Wczesną wiosną na roślinach wodnych pojawiają się intensywne brunatne naloty tworzone przez okrzemki, które później są zastępowane przez zielenice nitkowate (Tribonema, Spirogyra, Zygnema, Mougeotia, Oedogonium). Z początkiem lata kończy się zakwit glonów nitkowatych, a ich miejsce zajmują sinice (Microcystis, Aphanizomenon, Anabaena). Dla fitoplanktonu stawów typowe są także zielenice (Chlorella, Scenedesmus, Valvox), okrzemki (Melosira, Synedra) oraz rzadziej desmidie (Cosmarium, Staurastrum).
Plankton zwierzęcy budowany jest przede wszystkim przez wrotki (Conochilus, Keratella, Brachionus, Asplanchna, Synchaeta), liścionogi (Leptodora, Daphnia pulex, Daphnia longispina, Ceriodaphnia, Diaphanosoma, Bosmina) i widłonogi (Diaptomus, Cyclops). Ważną rolę w stawach odgrywa fauna poroślowa, czyli związana z roślinnością podwodną. Są to szeroko pojęte robaki, a szczególnie wrotki i skąposzczety, skorupiaki oraz larwy owadów. Natomiast w faunie dennej najważniejszą rolę odgrywają skąposzczety, przede wszystkim Tubificidae oraz larwy ochotkowatych, szczególnie czerwona larwa Chironomus plumosus.
Ze względu na cechy biologiczne Wurtz podzielił stawy na kilka kategorii:
- Stawy oligotroficzne:
- wykształcone na podłożu piaszczystym lub żwirowym, o wodzie obojętnej, dobrze natlenionej, ubogie produkcyjnie z różnorodną florą glonów, bez wyraźnej dominacji jakiejś grupy;
- wykształcone na podłożu piaszczystym lub gliniastym, o wodzie kwaśnej, słabo zamulonej, położone śródleśnie, z różnorodną florą glonów, bez wyraźnej dominacji;
- wykształcone na torfowiskach, z zalegającą na dnie słabo zmineralizowaną materią organiczną, dominują desmidie i okrzemki z Pennales.
- Stawy mezotroficzne - o słabo zamulonym dnie i dobrze natlenionej wodzie, nawet przy dnie; dominują chryzofity, obecne są bruzdnice i okrzemki, notowany często Botryocoocus.
- Stawy eutroficzne - o dobrze wykształconej warstwie mułu, przy powierzchni woda c zęsto przetleniona, natomiast przy dnie niedotleniona.
Dzielimy je na:
- słabo eutroficzne, z obfitą roślinnością naczyniową, na łodygach której licznie występuje peryfiton, dominują bruzdnice i okrzemki;
- średnio zeutrofizowane z obfitą roślinnością podwodną, w wodzie zasadowej dominują zielenice chlorokokkowe, zaś w kwaśnej bruzdnice, częste zakwity: wiosną - Volvox, latem - Dinobryon, jesienią - Microcystis;
- silnie zeutrofizowane z dominacją sinic (Anabaena) oraz liczną obecnością okrzemek, toczkowych i euglenin.
- Stawy politroficzne - z dużymi pokładami mułu na dnie i częstymi zanikami tlenu, dominuje sinica Aphanizomenon flos-aquae oraz toczkowe, chlorokokkowe i eugleniny.
Rzeki
Rzeki różnią się od zbiorników wód stojących ciągłym ruchem postępowym wody oraz kolejno ustawionymi za sobą biotopami. Stąd też oddziaływania między biotopami są jednokierunkowe, a nie wielokierunkowe tak jak w jeziorze. Powoduje to wykształcenie odmiennych niż w zbiornikach wody stojącej, układów biocenotycznych. Ważnymi elementami biocenotwórczymi są: duża zmienność linii brzegowej, niestabilność dna oraz wahania poziomu wody. Jest to szczególnie ważne w okresach wezbrań i powodzi, kiedy brzegi i dno są rozmywane, a osiadłe zbiorowiska roślin i zwierząt przestają istnieć. Natomiast w okresach suszy, przy znacznym obniżeniu lustra wody brzegowe biocenozy giną z przesuszenia.
W rzece wyróżnia się trzy odcinki charakteryzujące się swoistymi cechami siedliskowymi. Według Illiesa są to: krenon - odcinek źródłowy, rhitron - potokowy, potamon - rzeczny. Źródła są to miejsca, gdzie wody podziemne wydostają się na powierzchnię, dając początek potokom. Wody źródlane charakteryzują się małymi wahaniami temperatury w ciągu całego roku - latem są one zimne, zimą ciepłe i nie zamarzają. Zasiedlają je więc organizmy stenotermiczne. Pozostałe odcinki rzeki różnią się między sobą stopniem spadku, co determinuje charakter czynników fizycznych - siły prądu wody, rodzaju dna, zawartości tlenu, stopnia zanieczyszczenia, a w efekcie obecności określonych gatunków ryb na danym odcinku rzeki (rys. 35).
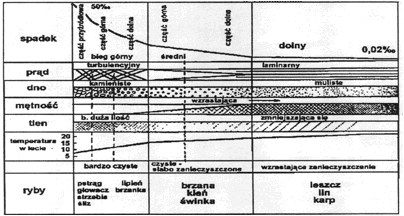
Rys. 35. Zmienność czynników środowiskowych wraz z biegiem rzeki
Źródła, czyli miejsca wypływu wody podziemnej na powierzchnię skorupy ziemskiej, mogą być z punktu widzenia geomorfologii, różnie wykształcone. Zależy to od układu warstw przepuszczalnych i nieprzepuszczalnych, a przede wszystkim od miejsca, w którym warstwa przepuszczalna wychodzi na powierzchnię. Thienemann wyróżnił trzy typy źródeł (rys. 36). Źródła reokrenowe, wywierzyskowe, charakteryzują się tym, że woda wypływa na zboczu i ma szybki nurt; dno takiego źródła jest czyste i pozbawione mułu. Źródło limnokrenowe to takie, w którym woda źródlana gromadzi się w nieckowatym zagłębieniu tworząc swego rodzaju zbiornik wodny. Woda z takiego źródła odpływa w jednym miejscu. Brzegi zbiornika źródlanego są zarośnięte mchami, wątrobowcami, niekiedy nawet roślinami kwiatowymi, zaś piaszczyste dno może być pokryte w niektórych miejscach warstewką mułu. Gdy warstwa wodonośna wychodzi na dużej przestrzeni, mamy do czynienia z tzw. źródłem helokrenowym, czyli bagiennym. Jest to więc zwykle podmokłe, zarośnięte bagnisko, na którym woda wysącza się i zgodnie z nachyleniem terenu spływa, tworząc w końcu strumień. Ten typ źródła jest właściwy dla rzek mających swój początek na niżu.
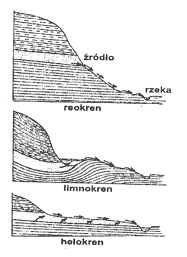
Rys. 36. Trzy typy źródeł.
Skład chemiczny wody źródła zależy od geologicznych właściwości jego podłoża. Tak więc zasobność w biogeny jest różna. Źródła helokrenowe są zwykle ubogie - rozwój roślinności jest tam słaby. Ubogie są również źródła reokrenowe. Najzasobniejsze w biogeny są źródła limnokrenowe.
Flora i fauna źródeł nie jest dokładnie zbadana. Spośród glonów występują w nich okrzemki (Meridion circulare, Diatoma hiemale var. mesodon i inne), zielenice (Draparnaldia, Tribonema) i krasnorosty (Batrachospermum). Ponadto obecne są mchy (Cratoneuron, Bryum, Philonotis) oraz - z roślin kwiatowych - rukiew wodna (Cardamine amara). Fauna reprezentowana jest przez drobne skąposzczety, skorupiaki, larwy różnych gatunków owadów.
W rzece obecne są w zasadzie wszystkie formacje ekologiczne, jednak niektóre z nich wykształcają się pełniej w wybranym odcinku jej biegu. Dwa czynniki środowiskowe zasadniczo określają typ zasiedlenia wód płynących. Jest to prąd i temperatura wody. Wartości tych czynników zmieniają się w sposób ciągły wzdłuż biegu rzeki, a zmienność ta jest taka sama we wszystkich rzekach, niezależnie od ich położenia geograficznego. Jakość warunków środowiskowych jest diametralnie różna w odcinku potokowym i rzecznym (rys. 35). Potok i rzeka różnią się bardzo wyraźnie pod względem warunków ekologicznych, wykształcają się w nich zasadniczo odmienne biocenozy. Rhitron, czyli odcinek potokowy, często określany jest jako kraina mikrofitów, względnie kraina ryb łososiowatych, zaś potamon, czyli odcinek rzeczny jako kraina makrofitów albo ryb karpiowatych.
Charakterystyczną cechą odcinka potokowego jest występowanie glonów o płaskich, skorupiastych plechach, przylegających do twardego, skalistego dna na odcinkach górskich. Są to przeważnie sinice (rząd Chamaesiphonales) lub krasnorost Hildenbrandtia rivularis. Liczne są także brunatne naloty okrzemek oraz bardzo charakterystyczny dla górnego biegu rzeki darnisto-nitkowaty złotowiciowiec Hydrurus foetidus. Ponadto obecnych jest kilka rodzajów zielenic (Gongrosira, Prasiola, Cladophora, Ulothrix, Draparnaldia, Oedogonium, Chaetophora), którym towarzyszą krasnorosty (Chantransia chalybaea, Lemanea torulosa, L.fluviatilis). Obok glonów w bystrych potokach żyją porosty (Verrucaria) i mchy wodne (Fontinalis, Cinclidotus) oraz rośliny kwiatowe (Potamogeton). Wśród glonów i innych roślin żyją liczne drobne zwierzęta - pierwotniaki, robaki, skorupiaki i larwy owadów, a przede wszystkim larwy jętek (Ephemeroptera), widelnic (Plecoptera), muchówek (Diptera), chruścików (Trichoptera). W bystrej wodzie obecne są na kamieniach wypławki (Planaria) i ślimak przytulik (Ancylus), zaś wśród gęstych zarośli gałęzatki (Cladophora) żyją kiełże (Gammarus) i rzadziej ośliczki (Asellus). Częste są także mocno przytwierdzone do kamieni gąbki i mszywioły. Obecne są także mikroskopijnych rozmiarów zwierzęta - wiciowce, wymoczki, wrotki, nicienie i drobne skorupiaki.
Odcinek rzeczny, czyli potamon, charakteryzuje się liczną obecnością dobrze rozwiniętych zbiorowisk roślin kwiatowych, ukorzenionych w dnie. Porastają one zwartymi łanami brzegi, opanowują płycizny oraz zajmują starorzecza. Towarzyszące im zespoły zwierząt są podobne do tych, jakie spotyka się w stawach i w strefie przybrzeżnej jezior. Obecne są więc ślimaki z rodzajów Limnea i Planorbis, oraz larwy ważek i jętek, chruściki, a przede wszystkim bardzo liczne są larwy muchówek, szczególnie ochotkowatych. Ponadto na dnie bardzo licznie występują skąposzczety. Liczebność fauny dennej w rzekach uzależniona jest od charakteru dna. Na podłożu piaszczystym może być do 100 osobników na 1 m2, natomiast w rejonach o grubej warstwie mułu zliczyć można do 2000 i więcej egzemplarzy w 1 m2 powierzchni dna.
W odcinkach niżowych wielkich rzek rozwija się plankton rzeczny, czyli potamoplankton. Charakteryzuje się on obfitością zawiesiny organicznej i mineralnej; jest jej zwykle więcej niż organizmów planktonowych. Zaznacza się przewaga ilościowa fitoplanktonu nad zooplanktonem. W fitoplanktonie najwięcej jest okrzemek, zaś w zooplanktonie wrotków. Ponadto w potamoplanktonie obecna jest niewielka ilość form typowo planktonowych, rozmnażających się w nurcie rzeki. Wykształcenie się potamoplanktonu jako formacji ekologicznej zależne jest od prędkości prądu wody rzecznej. Im prędkość jest mniejsza, tym lepsze są warunki do rozwoju planktonu. Szybkość przepływu 0,5-0,8 m/s pozwala już, jeśli rzeka jest odpowiednio długa, na rozwój gatunków planktonowych, powodujących nawet zakwity. Tak więc prawdziwy plankton nie rozwija się w pobliżu źródeł - woda źródlana musi przebyć kilkunastodniową drogę, aby zostały wytworzone odpowiednie warunki do rozwoju planktonu.
Skład nektonu wykazuje wyraźne zróżnicowanie w zależności od mierzonego w promilach spadku koryta rzeki. W rzekach o spadku poniżej 1o/oo panuje leszcz, gdy spadek wynosi 1-2o/oo - brzana, zaś powyżej 2o/oo - ryby łososiowate (lipień i pstrąg).
Woda rzeczna ostatecznie uchodzi do morza. W miejscu ujścia następuje mieszanie się niesionych przez rzekę wód słodkich ze słonymi wodami morskimi. Powoduje to wykształcenie się specyficznego obszaru wód słonawych tzw. estuarium. Z punktu widzenia ekologicznego jest to swoista strefa, o typowym charakterze ekotonu (czyli środowisko stykowe), gdzie bytują obok siebie gatunki typowe dla dwóch różnych ekosystemów. Cechą charakterystyczną estuarium jest to, iż w warstwie powierzchniowej rozpościerają się wody rzeczne, lżejsze od słonych wód morskich. Wciskane wskutek działania falowania wiatrowego (tzw. cofka) wody morskie zajmują warstwę przydenną. Tak więc w dostatecznie głębokim estuarium wykształcają się dwa obszary wód zróżnicowanych pod względem zasolenia, co determinuje obecność odpowiednich grup organizmów w przekroju pionowym. W górnych warstwach przeważają formy potamoplanktonu, zaś w dolnych morskie. Prócz tego w rejonie estuarium obecnych jest wiele gatunków euryhalinowych. Różnorodność gatunkowa w obszarach ujść rzecznych jest więc duża - są to na ogół bogate rejony rybackie.
Produktywność ekosystemów wodnych
Wytworzona w ekosystemie biomasa podlega, poprzez wzajemne relacje między organizmami, różnym przemianom. Jest ona zjadana, przyswajana i stanowi budulec biomasy innych organizmów. W przemianach tych część jej zostaje zużyta na procesy metaboliczne i po obumarciu organizmu ulega rozkładowi bakteryjnemu, a niekiedy jako nie do końca rozłożona materia organiczna może zalegać na dnie w osadach. Tak więc podstawowa zależność w ekosystemie ma charakter pokarmowy, czyli troficzny. Organizmy autotroficzne, jako producenci materii organicznej, stanowią pierwszy poziom troficzny. W zbiorniku wodnym ten poziom stanowi fitoplankton, makrofity, mikrofitobentos i epifity. Na nich żerują konsumenci kolejnych poziomów troficznych. Konsumentami I rzędu są: zooplankton roślinożerny, bakterie i fauna denna roślinożerna. Grupa ta stanowi pokarm dla konsumentów II rzędu: zooplanktonu mięsożernego i fauny dennej mięsożernej. Następny poziom konsumentów stanowią ryby planktonożerne oraz ryby żerujące na drobnej faunie dennej. Ostatnie ogniwo stanowią drapieżne ryby i inni drapieżnicy. Układ ten przedstawiony w formie łańcucha troficznego w warunkach naturalnych jest bardziej skomplikowany i powinien być przedstawiony w postaci sieci troficznej. Podstawą zależności troficznych jest produkcja materii organicznej.
Produkcja biologiczna w zbiorniku wodnym potocznie kojarzy się z ilością ryb, które można wyłowić i skonsumować. Aby jednak ten końcowy efekt mógł zaistnieć musi przede wszystkim nastąpić przetworzenie składników nieorganicznych i energii promienistej w procesie fotosyntezy w materię organiczną. Proces ten określany jest nazwą produkcji podstawowej lub pierwotnej. Zmierzenie ilości wytworzonych w tym procesie związków organicznych węgla przez rośliny jest dość proste, chociażby poprzez określenie ilości związanego podczas fotosyntezy dwutlenku węgla lub równoważnie wydzielonego tlenu. Mając te dane i stosując odpowiednie przeliczenia, można określić wielkość biomasy wyprodukowanej przez konsumentów. Jest to tzw. produkcja wtórna. Oprócz producentów i konsumentów trzecim ogniwem każdego ekosystemu są heterotroficzne mikroorganizmy tzw. reducenci. Redukują one materię organiczną do postaci prostych związków nieorganicznych, dzięki czemu może istnieć w zbiorniku wodnym obieg materii. Produkcja biologiczna jest więc procesem, w którym zostaje wytworzona odpowiednia ilość materii organicznej na danej przestrzeni w określonym czasie. Może to być przyrost biomasy w ciągu dnia, miesiąca albo roku. W przypadku bentosu odnosi się ją do jednostki powierzchni (np. 1 m2), a dla innych formacji ekologicznych do jednostki objętości (np. 1 m3 wody). Ponieważ w procesie fotosyntezy wiązana jest energia promienista, która jako energia chemiczna z pokarmem jest przenoszona na kolejne poziomy troficzne to można mówić o przepływie energii przez ekosystem. Przyjmuje się, że jedynie 2% energii świetlnej jest wiązane w procesie produkcji pierwotnej. Natomiast przy przejściu na każdy następny poziom troficzny starty sięgają 90%. Straty te związane są z metabolizmem, w którym oddychanie odgrywa podstawową rolę. W procesie tym energia chemiczna z pokarmu zostaje przetworzona w energię cieplną i podlega entropii, czyli rozproszeniu i jest ona dla procesów ekosystemowych nieużyteczna. Przepływ energii jest więc zjawiskiem jednokierunkowym. Uwzględniając procesy metaboliczne wyróżnia się dwa rodzaje produkcji. Pierwsza z nich to produkcja ogólna (brutto), czyli ta ilość energii, która zostaje ogólnie pobrana i przerobiona. Druga to produkcja czysta (netto), a więc ilość energii pobranej i przerobionej, czyli produkcja brutto zmniejszona o straty energii związane z procesem oddychania oraz energię wydaloną w postaci odchodów, moczu itp.
Produktywność zbiornika, czyli jego wydajność biologiczną, określa się przez potencjał produkcyjny, a więc sprawność przyrostu biomasy. Produktywność jest efektem produkcyjnym danego zbiornika, mówi się zatem o dużej lub małej produktywności wybranych jezior, rzek lub mórz. Najczęściej ten potencjał produkcyjny przedstawiany jest w postaci wielkości produkcji pierwotnej, a to ze względu na dość dużą łatwość eksperymentalnego pomiaru. Do tego celu służy kilka metod, z których najczęściej stosowane są metody: tlenowa i węgla radioaktywnego.
Metoda tlenowa zastosowana w latach dwudziestych do badań produkcji planktonu w morzach, przystosowana została w latach trzydziestych do badań w wodach słodkich. Polega ona na ekspozycji przez określony czas (np. dobę) na danej głębokości dwóch butelek z wodą pochodzącą z tej głębokości. Jedna z butelek jest umieszczona w osłonie ciemnej (np. pomalowana czarną farbą) - dlatego właśnie pomiar ten często określany jest jako metoda jasnych i ciemnych butelek. Przed ekspozycją i po jej zakończeniu mierzy się w obu butelkach zawartość tlenu metodą Winklera. Z wyliczonej różnicy zawartości tlenu przed i po ekspozycji w jasnej butelce określa się wielkość produkcji pierwotnej całkowitej (brutto). Po uwzględnieniu zużycia tlenu na oddychanie zmierzonego w ciemnej butelce uzyskuje się wielkość produkcji pierwotnej czystej (netto). Uzyskane wielkości wyprodukowanego tlenu można przeliczyć za pomocą równania fotosyntezy na ilość zużytego dwutlenku węgla lub ilość kalorii uzyskanych w tym procesie. Czułość tej metody jest ograniczona. Stosować ją można, gdy wielkość produkcji wynosi powyżej 10 mg C/m3. Jest więc ona zawodna w badaniach produkcji pierwotnej jezior oligotroficznych. Metoda ta ma także kilka innych mankamentów obniżających jej czułość. I tak wyniki zaniżone daje zbyt długa ekspozycja i również sfalowana powierzchnia wody. Ponadto na wynik wpływa obecność w butelkach bakterii i planktonu zwierzęcego.
Metoda węgla radioaktywnego C14 jest czulsza i umożliwia dokładniejszy pomiar, tj. do 0,1 mg C/m3 wody. Po raz pierwszy została zastosowana w 1952 roku przez Duńczyka Steemanna-Nielsena w badaniach morskich, a potem znalazła powszechne zastosowanie także w badaniach słodkowodnych. Polega ona na wprowadzaniu do butelek, służących do ekspozycji, znakowanego kwaśnego węglanu sodu. Węglan ten zostaje pobrany przez glony w trakcie fotosyntezy. Po ekspozycji zawartość butelki przesącza się i wylicza radioaktywność próby w liczniku Geigera. Metoda ta jest również obarczona zastrzeżeniami o charakterze technicznym (czas ekspozycji, rodzaj szkła butelek używanych do ekspozycji) i środowiskowym (temperatura wody, typ glonów, obecność zooplanktonu w próbie).
Na podstawie dotychczasowych badań uważa się, że produkcja pierwotna różnych typów wód mieści się w granicach 0,01-24 g C/m2 na dobę. Różnice są więc bardzo duże i zależą przede wszystkim od strefy klimatycznej i zasobności wody w substancje mineralne. Jeziora arktyczne dają produkcję pierwotną w granicach 0,03-21,3 mg C/m2 na dobę. Jeziora oligotroficzne strefy umiarkowanej charakteryzują się również niską produkcją wynoszącą około 100 mg C/m2 na dobę. Jeziora eutroficzne wykazują produkcję pierwotną około 200-2000 mg C/m2 na dobę. Rzeki, ze względu na warunki środowiskowe (niestabilny biotop), dają znacznie mniejsze wartości. Ze względu na ogrom obszaru mórz i oceanów w stosunku do zbiorników śródlądowych, produkcja pierwotna w tych ostatnich stanowi znikomy odsetek całości tej produkcji.
W procesach produkcyjnych oceanów podstawową rolę odgrywa fitoplankton stanowiący ponad 90% zasobów pożywienia zwierząt morskich. Rejonizacja produkcji pierwotnej w oceanach jest bardzo zbliżona do rozmieszczenia ilościowego fitoplanktonu. Najbardziej produktywne rejony to te, gdzie najszybciej zachodzi odnawianie biogenów. Są więc to obszary przybrzeżnego wynoszenia wód (upwelling) i dywergencji prądów. Dlatego też obraz produkcji pierwotnej w oceanach odzwierciedla dwa typy strefowości: równoleżnikowej i wzdłużkontynentalnej. W Oceanie Atlantyckim znaczna większość wód odznacza się umiarkowaną produkcją pierwotną wynoszącą średnio 100-250 mg C/m2 na dobę. Zróżnicowanie tej wartości jest jednak dość znaczne i waha się od 0,02-0,03 g C/m2 na dobę w Morzu Sargassowym do 3-5 g C/m2 na dobę u wybrzeży Namibii. W Oceanie Spokojnym wielkość produkcji pierwotnej waha się od kilkudziesięciu miligramów do 650 mg węgla pod 1 m2 na dobę. Najmniejszą produkcją charakteryzują się rejony subtropikalne i tropikalne. Największe wartości produkcji, sięgające 500 mg C/m2 na dobę, notowane są w północnych wodach krainy borealnej, przy brzegach Kamczatki, Japonii i Alaski. Najbardziej produktywny jest Ocean Indyjski, gdzie średnia wielkość produkcji pierwotnej wynosi 222 mg C/m2 na dobę, podczas gdy dla Oceanu Atlantyckiego - 190 mg C/m2 na dobę, a Oceanu Spokojnego - 127 mg C/m2 na dobę. Globalna produkcja pierwotna wód jest o około 2,5 raza mniejsza od produkcji szaty roślinnej na lądzie, mimo iż wody zajmują 2/3 powierzchni kuli ziemskiej. Wynika to z lepszego wykorzystania energii świetlnej na lądzie.
Sprawność produkcyjna ekosystemu, czyli jego produktywność końcowa, a więc wielkość tzw. plonu, uzależniona jest od gospodarowania energią zmagazynowaną na etapie produkcji pierwotnej. Przepływ energii, czyli jej droga wzdłuż łańcucha troficznego, związany jest ściśle z poszczególnymi składnikami ekosystemu. Natężenie przepływu energii zależy od struktury ekologicznej zbiornika, inne więc będzie w rzece jako ekosystemie otwartym, inne w jeziorze jako ekosystemie zamkniętym. Intensywniejszy jest przepływ energii w ekosystemach otwartych. Badania nad przepływem energii w ekosystemie muszą być całościowe. Konieczne jest więc szczegółowe rozpoznanie zasobów energetycznych na wszystkich poziomach troficznych w odniesieniu do producentów, konsumentów i reducentów. Poznanie jedynie energii wyjściowej i jej stanu na jednym poziomie nie pozwala na wyciąganie wniosków o całości ekosystemu.
Problemy hydrobiologii stosowanej
Eutrofizacja
Przez pojęcie eutrofizacji rozumie się proces wzbogacania wód w mineralne sole biogeniczne, tak na drodze naturalnej, jak i sztucznej. Właśnie w takim znaczeniu, na określenie wzbogacenia wód jeziornych, użył tej nazwy Hasler w 1947 roku. Pod tym pojęciem rozumie się więc proces dynamicznej zmiany stanów (oligotroficzny - eutroficzny). Eutrofizacja w zbiorniku wodnym objawia się zwiększoną produkcją materii organicznej. Przyczyny tego zjawiska są różne: warunkują je czynniki chemiczne, biologiczne, fizyczne i natury geograficznej związane z klimatem. Tak więc eutrofizacja zależy od całego kompleksu czynników i jest zjawiskiem stałym, związanym ze spływem wód opadowych ze zlewni. Stąd też zachodzi on tak długo, jak długo istnieją zbiorniki wodne. Zjawisko takie nazywano eutrofizacją naturalną ponieważ przyczyny jego mają pochodzenie naturalne (przyrodnicza erozja zlewni).
Naturalny przebieg procesu eutrofizacji został zintensyfikowany działalnością człowieka, a przede wszystkim spływem różnego rodzaju ścieków (komunalnych, przemysłowych, rolniczych). Istotną rolę gra także np. zwiększenie areału ziemi uprawnej powodujące zmniejszenie powierzchni lasów. Wpływa to na zmianę warunków hydrologicznych zlewni i przyspieszenie procesów erozyjnych. Zwiększony zostaje w ten sposób spływ materiału ze zlewni i w efekcie następuje wypłycenie zbiorników wodnych. Naniesiona materia organiczna i zmyte mineralne związki biogeniczne powodują wzrost produkcji biologicznej. Przyspieszenie procesu eutrofizacji wywołane przez działalność człowieka zostało określone nazwą eutrofizacji sztucznej. Jest ona nieodłącznym atrybutem cywilizacji i związane jest to z przekształcaniem przez człowieka otoczenia.
Związek pomiędzy procesem eutrofizacji wód a intensyfikacją nawożenia upraw jest oczywisty. Spośród wielu pierwiastków potrzebnych do rozwoju roślin najważniejszymi są: węgiel, azot i fosfor. One są też podstawową przyczyną sprawczą eutrofizacji. O ich decydującej roli można przekonać się porównując zapasy węgla, azotu i fosforu w wodach naturalnych i ściekach komunalnych z zapotrzebowaniem do produkcji biologicznej wynikającym ze stosunku C:N:P w ciele glonów wynoszącym 40:7:1. W wodach naturalnych zapasy węgla są w nadmiarze, np. w zbiornikach oligotroficznych stosunek C:N:P = 600:20:1. Zupełnie inaczej stosunek ten kształtuje się w wodach zanieczyszczonych, np. w zmineralizowanych ściekach komunalnych wynosi 6:4:1. Wynika stąd, że pierwiastkami limitującymi produkcję i decydującymi o procesie eutrofizacji są azot, a szczególnie fosfor. Zasadnicza rola tego ostatniego pierwiastka wynika także z łatwiejszej dostępności azotu - w wielu typach wód, szczególnie w warstwie powierzchniowej, azot może być uzupełniany z powietrza dzięki zdolnościom fiksacji wolnego azotu przez sinice. W takiej sytuacji proces eutrofizacji jest silnie uzależniony od stężenia fosforanów w wodzie.
Źródła soli biogennych można podzielić na trzy grupy: ścieki przemysłowe, spływ komunalny i z gospodarstw wiejskich oraz spływ ze zlewni (ługowanie i erozja gleb). Ważną rolę odgrywają również opady atmosferyczne. Duża ilość związków azotowych, niekiedy ponad 50% całości, pochodzi z ługowania gleb. Głównym źródłem fosforu są wody odpadowe z gospodarstw domowych, w czym istotną rolę odgrywają środki piorące (detergenty). Duże ilości soli biogennych zawarte są w ściekach z zakładów przemysłu rolnego (krochmalnie, cukrownie, rzeźnie, gorzelnie, mleczarnie, browary, drożdżownie). Mogą one stanowić 50-70% ładunku zawartego w ściekach miejskich.
Znaczne ilości biogenów, szczególnie związków azotowych, dostają się z wód gruntowych do wód powierzchniowych w wyniku przemywania gleb. Związane jest to, o czym już wspomniano wyżej, z gospodarką rolną. Z analizy osadów dennych jezior wynika, że z chwilą rozwoju rolnictwa w zlewni jeziora następuje gromadzenie się w osadach związków fosforu, potasu, żelaza i innych, a przede wszystkim materii organicznej. Współczesne nawożenie mineralne proces ten przyspiesza. Tak więc do zbiorników wodnych dostają się te związki chemiczne, które bezpośrednio powodują eutrofizację naturalną. Z gleb najintensywniej wypłukiwane są związki wapnia i azotu. Wapń wymywany jest w postaci rozpuszczalnych kwaśnych węglanów. Przenoszenie ze zlewni do zbiornika tej postaci związków wapnia, jak również magnezu, powoduje wzbogacenie wody w dwutlenek węgla.
Azot jest wymywany z gleb głównie w postaci azotanów. W rolnictwie stosuje się obecnie nawozy zawierające więcej formy amonowej, np. mocznik, saletra amonowa i siarczan amonowy. Nie ogranicza to jednak ilości azotanów w glebie ze względu na zachodzącą szybko hydrolizę mocznika oraz intensywną nitryfikację formy amonowej. W efekcie prowadzi to do wzrostu koncentracji azotu azotanowego, który nie jest zatrzymywany przez kompleks sorpcyjny gleb. Wymywanie azotu z gleb uzależnione jest od poziomu nawożenia, płodozmianu uprawy i właściwości mechanicznych gleb, szczególnie ich przepuszczalności. Gleby lekkie, przepuszczalne, których w Polsce jest ponad 50%, sprzyjają nitryfikacji, a w efekcie wymywaniu azotanów. Zjawisko wymywania związków azotowych jest zwielokrotnione w okresie wiosennych powodzi, kiedy to równocześnie od wysiewu nawozów mineralnych zaczynają się roboty polowe. Przyjmuje się, że odpływ azotu z gleb uprawnych w Polsce wynosi 10-20 kg/ha rocznie.
Związki fosforu i potasu dzięki właściwościom sorpcyjnym gleb są wymywane w znacznie mniejszym stopniu niż nawozy azotowe. Stąd też wymywanie fosforu jest niewielkie i zasadniczo nie przekracza 0,5 kg/ha rocznie. Fosforany, stosowane szeroko w postaci superfosfatu, szybko wiązane są chemicznie w glebie i dlatego przemieszczanie ich jest bardzo powolne. Podobnie sole potasowe i amonowe są wiązane w kompleksie sorpcyjnym gleb, jednak wiązanie to słabnie w miarę wzrostu kwasoty gleby. Staje się to coraz bardziej poważnym problemem wywołanym rozszerzającym się zjawiskiem acydyfikacji, czyli zakwaszania środowiska naturalnego.
Rola opadów atmosferycznych jako źródła biogenów zależy od stopnia zanieczyszczenia powietrza. W opadach tych występuje stosunkowo dużo związków azotowych, głównie amonowych. Przewaga formy amonowej nad azotanową związana jest ze stopniem zanieczyszczenia powietrza. Szacuje się, że ilości związków azotowych pochodzących z opadów atmosferycznych wynoszą rocznie w rejonach rolniczych około 10 kg/ha, a w przemysłowych ponad 20 kg/ha, zaś fosforu tą drogą dostaje się około 0,5 kg/ha.
Walka z eutrofizacją polega na ograniczeniu przyczyn nadmiernego wzrostu roślin wodnych, w tym przede wszystkim fitoplanktonu, oraz likwidacji skutków tego zjawiska. Ograniczenie przyczyn eutrofizacji polega na eliminowaniu soli biogennych poprzez oczyszczanie ścieków. Dość łatwo dają się eliminować fosforany, poprzez wytrącanie ich w nierozpuszczalnych solach żelaza, glinu lub wapnia. Na przykład dodanie do ścieków w trakcie ich oczyszczania chlorku żelazowego eliminuje około 90% fosforanów. Metod podobnych nie można stosować do eliminacji ani azotanów, ani soli amonowych, gdyż są one dobrze rozpuszczalne w wodzie. Próbuje się w tym przypadku stosować metody fizykochemiczne polegające na alkalizowaniu ścieków i usuwaniu gazowego amoniaku lub stosowaniu wymieniaczy jonowych. Metody te mają wiele mankamentów i są mało wydajne. Większą wydajnością charakteryzują się metody biologiczne oparte na denitryfikacji azotanów. Duże nadzieje budzą metody, w których glony są użyte jako eliminatory biogenów. Jest to typowy przykład oczyszczania biologicznego, dzięki któremu można by uzyskać prawie zamknięty obieg biogenów, gdyż zmagazynowane metabolicznie w glonach biogeny, pochodzące ze ścieków, mogą być stosowane w rolnictwie jako doskonały nawóz.
Przedstawione metody walki z eutrofizacją dotyczą tzw. punktowych źródeł zanieczyszczeń. Sytuacja jest znacznie bardziej skomplikowana w przypadku rolniczych źródeł eutrofizacji, gdyż mamy tu do czynienia z przestrzennym zanieczyszczeniem. Eutrofizację rolniczą można ograniczyć przez stosowanie nawozów wiązanych w glebie, tym samym wolniej z niej wymywanych i pełniej wykorzystywanych przez rośliny uprawne. Ważne są w tym przypadku także zabiegi mające na celu zwiększanie kompleksu sorpcyjnego i ograniczenie erozji gleb.
Saprobizacja
Nazwą tą można określić zjawisko zanieczyszczania wód naturalnych w wyniku działalności gospodarczej. Jest ono przedmiotem badań nauki zwanej saprobiologią lub biologią ścieków. Zadaniem tej nauki, wspólnie z odpowiednimi działami techniki, jest poznanie, ograniczenie i usuwanie zanieczyszczeń. Zanieczyszczenie wód naturalnych jest jednym z efektów rozwoju cywilizacji. Ścieki, czyli woda przez człowieka zużyta, są nieodzownym elementem ludzkiej działalności. O wodach ściekowych i higienie życia codziennego wspominają najstarsze zapisy cywilizacyjne. Jednak nowoczesne badania biologiczne wód ściekowych datują się dopiero od połowy ubiegłego stulecia. Saprobiologia jest nauką szczególnie intensywnie rozwijającą się w krajach znacznie zurbanizowanych i wysoko uprzemysłowionych.
Dość trudno jest postawić jednoznaczną granicę między wodą zeutrofizowaną a zanieczyszczoną. Definicja wody zanieczyszczonej jest nieostra. Przyjmuje się, że woda zanieczyszczona to taka, której stan jest tak zmieniony, iż użytkowanie jej do celów domowych i przemysłowych jest ograniczone lub niemożliwe. Wody silnie obciążone biogenami w stanie tzw. politrofii lub nawet hipertrofii, mogą funkcjonować według normalnych prawideł obiegu materii. Jednak sytuacja, w której następuje taka kumulacja materii organicznej, że procesy obiegu materii są zahamowane, przypomina stosunki panujące w zbiornikach nieharmonicznych. Stan ten określa się nazwą saprotrofii. Tak więc zbiornik saprotroficzny to taki, w którym procesy destrukcyjne, czyli rozpadowe (czasami określane jako stan saprobii), przeważają nad procesami produkcyjnymi (czyli nad trofią), mimo dużej zawartości biogenów w formie wolnej lub związanej w materii organicznej.
Ściek jest to dopływ wnoszący do wody naturalnej substancje zanieczyszczające. Wyróżnia się:
- ścieki gospodarcze, inaczej domowe, powstałe w wyniku używania wody do celów domowych, obciążone są one w znacznym stopniu odchodami;
- ścieki komunalne, pochodzące z osiedli, zawierają one ścieki gospodarcze i przemysłu komunalnego, z mleczarni, rzeźni itp.;
- ścieki przemysłowe, z dużych zakładów chemicznych, celulozowych, garbarskich i in.
Ścieki w chwili wpływania do odbieralnika (odbieralnik to zbiornik wodny, do którego spływają ścieki), ulegają rozcieńczeniu i zaczyna się proces samooczyszczania. Samooczyszczanie się wody jest procesem biologicznym. Polega on na adsorpcji i resorpcji przez organizmy żywe zanieczyszczeń oraz na rozkładzie i mineralizacji rozpuszczonej materii organicznej przy udziale mikroorganizmów. Ścieki przemysłowe zawierające cyjanki, fenole lub sole metali ciężkich powodują zatrucie organizmów, co wyklucza zaistnienie zjawiska samooczyszczania odbywającego się - jak powiedziano wyżej - dzięki procesom biologicznym.
Na pierwszym etapie samooczyszczania aktywne są przede wszystkim te organizmy, które potrzebują materii organicznej jako pożywienia. Są to bakterie i grzyby, żywiące się materią organiczną. Do grupy tej należą też miksotroficzne glony, częściowo wykorzystujące materię organiczną rozpuszczoną w wodzie. Szybko namnażające się bakterie są usuwane ze środowiska, przez zjadające je zwierzęta - ameby, orzęski, wrotki, rozwielitki, skąposzczety, larwy owadów i inne. Tak więc na materii organicznej ścieków powstaje nowe życie. Przyjmuje się, że rzeka potrafi się sama oczyścić, gdy ilość ścieków organicznych w stosunku do czystej wody wynosi 1:50.
Ponieważ warunki życia i jego przejawy są przyczynowo powiązane, można na podstawie obecności organizmów żywych mówić o warunkach życia w danej wodzie. Na podstawie tych założeń wprowadzono system wskaźników biologicznych stopnia zanieczyszczenia. System ten sformułowany został w latach 1908-1910 przez Kolkwitza i Marssona i nazwany systemem saprobów.
System saprobów oparty jest na różnicach w składzie gatunkowym organizmów zasiedlających rzekę od wpłynięcia do niej ścieków, aż po ich oczyszczenie i zmineralizowanie. Organizmy, które zasiedlają to zanieczyszczone środowisko noszą nazwę saprobów (od greckiego słowa sapros - gnijący). W systemie Kolkwitza i Marssona wydzielono 4 grupy saprobów:
- polisaproby żyjące w wodach silnie zanieczyszczonych;
- α-mezosaproby - w wodach średnio zanieczyszczonych;
- β-mezosaproby - w wodach mniej zanieczyszczonych;
- oligosaproby - w wodach mało zanieczyszczonych.
Organizmy bytujące w wodach nie zanieczyszczonych określa się nazwą kataroby.
System saprobów opiera się na zasadzie analitycznego odnajdywania gatunków wskaźnikowych. Rozwijające się badania nad autekologią poszczególnych gatunków wskazały na ważny element ich gromadnego występowania. Dlatego w latach sześćdziesiątych Fjerdingstad zaproponował nowy system saprobów, oparty na zespołach organizmów. Późniejsze badania wykazały, że w związku z rozwojem przemysłu pojawiły się nowe typy ścieków, o specyficznym charakterze ekologicznym. Przyczyniło się to do nowego ujęcia systemu saprobów, w którym dla określenia zanieczyszczenia wód, obok saprobowości, wprowadzono stany toksyczności i radioaktywności, a dla określenia odpowiednich stopni zanieczyszczenia posłużyły zespoły gatunków proponowane przez Fjerdingstada oraz liczebność bakterii, a przede wszystkim zjawiska metaboliczne określające potencjał produkcyjny. W takim ujęciu system saprobów proponowany przez Sládečka obejmuje 4 kategorie wód:
- katarobowe najczystsze, zdatne do picia bez dodatkowego oczyszczania;
- limnosaprobowe - zanieczyszczone, dzielące się na: ksenosaprobowe, oligosaprobowe, α-mezosaprobowe, β-mezosaprobowe i polisaprobowe;
- eusaprobowe - ściekowe, obejmujące: izosaprobowe, hipersaprobowe i ultrasaprobowe;
- transsaprobowe - ścieki nie podlegające rozkładowi w procesach biologicznych, dzielące się na: antysaprobowe, kryptosaprobowe i radiosaprobowe.
Użytkowy aspekt badań saprobiologicznych został wykorzystany praktycznie przy technicznym oczyszczaniu ścieków w specjalnych urządzeniach, tzw. oczyszczalniach, szczególnie w procesie oczyszczania biologicznego. Pierwszym etapem oczyszczania ścieków jest oczyszczanie mechaniczne, w trakcie którego dzięki odpowiednim urządzeniom technicznym i seriom odstojników usuwane są większe materiały, zawiesina i substancje olejowe. Natomiast wśród biologicznych sposobów oczyszczania ścieków wyróżnia się zabiegi naturalne, półnaturalne i techniczne.
Zabiegi naturalne polegają na podlewaniu wodami ściekowymi łąk lub pól uprawnych. Wówczas obecne w glebie mikroorganizmy spełniają rolę reducentów mineralizujących materię organiczną zawartą w ściekach. Takiej utylizacji mogą podlegać tylko ścieki nietoksyczne. Metody półnaturalne to takie, kiedy ścieki przepływają przez kilka sztucznych zbiorników dobrze naświetlonych i natlenionych. Zachodzi w nich proces samooczyszczenia drogą naturalną. W końcowych etapach tego procesu zbiorniki te mogą być wykorzystane do hodowli ryb. Metody techniczne mają za zadanie uefektywnienie i przyspieszenie procesu samooczyszczania. W tym celu stosuje się tzw. osad czynny lub tzw. złoże biologiczne. Metody te charakteryzują się dużą wydajnością. Osad czynny to zawiesina ścieków z odpowiednimi grupami bakterii, najczęściej w postaci kłaczkowatych, śluzowatych skupień. Jest on stale mieszany i przewietrzany. Osad czynny powoduje skupianie się zawiesiny i koloidów, przy jednoczesnym rozkładzie chemicznym. Złoża biologiczne są to kilkumetrowej długości, cylindryczne kolumny wypełnione porowatym gruzem mineralnym - koksem lub żużlem. Od góry sączą się przez nie ścieki, zaś powietrze jest tłoczone pod ciśnieniem od dołu, tak więc w górnej części zachodzą procesy beztlenowe, natomiast w dolnej tlenowe, z przewagą nitryfikacji, dzięki obecnym tam bakteriom, pierwotniakom, wrotkom, nicieniom, a nawet larwom owadów. Oczyszczanie biologiczne nie usuwa soli biogennych ze ścieków, dlatego też ważny jest etap oczyszczania chemicznego - chemicznie nie oczyszczone wody pościekowe powodują eutrofizację.
Acydyfikacja
Terminem tym przyjęto określać zjawisko zakwaszania środowiska. Jego przyczyną jest obecność w atmosferze dwutlenku siarki i tlenków azotu. Wskutek intensyfikacji przemysłu i motoryzacji występują one w nadmiernych ilościach. W niedużych ilościach tlenki siarki i azotu występują normalnie w atmosferze. Deszcze, bez wpływu działalności człowieka, powinny mieć odczyn w granicach pH=5-6 (z lokalnymi wahaniami). Według najnowszych danych, na szerokich przestrzeniach Europy pH wody deszczowej jest dużo mniejsze, między 4,5 i 5. W pojedynczych przypadkach notowano jeszcze mniejsze wynoszące około 3. Malejące pH jest wskaźnikiem wrastającej kwasowości; gdy pH obniża się o jedną jednostkę, to stężenie kwasu (liczba jonów wodorowych) wzrasta ponad dziesięciokrotnie.
Część emitowanego z kompleksów przemysłowo-komunalnych dwutlenku siarki (S02) i tlenku azotu (NO) opada na ziemię w pobliżu - mówimy wówczas o tzw. suchym opadzie. Pozostała, większa część jest przenoszona na znaczne odległości ruchami powietrza. W tym samym czasie następuje ich utlenianie i tworzy się kwas siarkowy (H2SO4) i kwas azotowy (HNO3), które w wilgotnym środowisku chmur ulegają dysocjacji. W takiej formie mieszanki jonów: SO4--, NO3- i H+ dostają się na powierzchnię ziemi podczas opadów. Jest to tzw. opad mokry w formie "kwaśnych deszczy". Kumulacja tych jonów zachodzi w glebie i w zbiornikach wody naturalnej. Opad suchy po spadnięciu na ziemię, a w szczególności do wody, ulega transformacji jonowej i daje w efekcie kwasy. W związku z możliwością przenoszenia tlenków w powietrzu na znaczne odległości, mogą one spadać na ziemię jako mokry opad w odległości setek bądź tysięcy kilometrów od źródła emisji.
W polskich opadach atmosferycznych notuje się dużą ilość związków siarki. Spowodowane jest to właśnie zanieczyszczeniem atmosfery dwutlenkiem siarki. Roczny opad tych związków w Polsce w rejonach rolniczych waha się w granicach 10-15 kg/ha, a w okręgach przemysłowych 32-95 kg/ha.
Proces acydyfikacji zbiorników wodnych przebiega zasadniczo w trzech etapach. Gdy skały zlewni i otaczające zbiornik gleby są łatwo rozpuszczalne, woda z nich spływająca będzie już zobojętniona. Zawartość dwuwęglanu w takiej wodzie będzie duża. Przypadkowy wzrost stężenia kwasów (wysoka zawartość jonów wodorowych) będzie zobojętniany przez nadwyżki jonów dwuwęglanowych (podczas reakcji jonów wodorowych z jonami dwuwęglanów tworzy się dwutlenek węgla i woda). W tym okresie (pierwszy etap) nie stwierdza się spadku pH, jednak zawartość jonów węglanowych po pewnym czasie zacznie spadać. Konsekwencją tego będzie stopniowe osłabienie zdolności buforującej, czyli zobojętniania dopływających kwasów i w pewnym momencie, gdy dopływ nie ustanie, zasoby dwuwęglanu zostaną wyczerpane. Podczas tego pierwszego etapu acydyfikacji nie stwierdza się zmian w procesach życiowych w zbiornikach wodnych.
Gdy zawartość dwuwęglanów spada tak bardzo, że dalej dopływające jony wodorowe nie są już neutralizowane, zaczyna się drugi etap acydyfikacji. Wartość pH staje się niestabilna i zaczyna spadać znacznie szybciej niż poprzednio. Zaczyna się zaznaczać ujemny wpływ zmiany pH na życie w wodzie. Równowagę zakłócają silne jesienne deszcze obniżające pH wody - w tym okresie acydyfikacji notuje się znaczne wahania pH w cyklu rocznym. Ma to swoje odbicie w bezpośrednich i rozległych biologicznych szkodach, takich jak masowe śnięcie ryb lub spadek zdolności reprodukcyjnej wielu gatunków ryb. W tym etapie acydyfikacji wartość pH wód powierzchniowych w niektórych okresach roku jest mniejsza od 5,5. Jeżeli dopływ kwasów się nie zwiększy, etap drugi zakwaszenia może się cofnąć, natomiast w przeciwnym razie acydyfikacja przyjmuje bardzo poważny obrót.
W trzecim etapie acydyfikacji wartość pH wody w zbiorniku stabilizuje się wokół 4,5 nawet jeśli opad jest bardziej kwaśny, a dopływ jonów wodorowych stały. Na tym etapie zakwaszenia substancje humusowe i glin zaczynają działać jako bufory. Substancje humusowe, które co prawda są kwasami, "wyłapują" jony wodorowe zamiast uwalniać je do środowiska. W średnio kwaśnych wodach jony glinu (Al+++) funkcjonują jako kwas i reagują z wodą tworząc wodorotlenek glinu. Dalsze zakwaszanie powoduje rozpuszczanie Al(OH)3, co w efekcie neutralizuje pH (Al(OH)3 + 3H+ --> Al+++ + 3H2O). W kwaśnych wodach powierzchniowych drastycznie wzrasta zawartość jonów glinu. Ma to silnie toksyczny wpływ na wiele organizmów. Tak więc oczywiste jest, że rzeczywistą przyczyną masowych śnięć ryb są zatrucia aluminium. W trzecim stadium acydyfikacji mamy do czynienia z całkiem nowym ekosystemem, gdzie fauna ryb w większości znikła, a liczba dominujących gatunków roślin jest mała. Cechą charakterystyczną wielu zacydyfikowanych jezior jest duża przezroczystość wody, która zwiększa się z około 4-5 m w jeziorach normalnych do 15-20 m w zakwaszonych.
Oprócz wzrostu zawartości glinu, w zakwaszonej wodzie zachodzą również inne reakcje chemiczne. Wzrastająca rozpuszczalność toksycznych metali ciężkich (kadm, cynk, ołów, magnez i inne), ma swoje odbicie w procesach życiowych roślin i zwierząt. Spada zawartość niektórych biogenów w zakwaszonej wodzie. W zacydyfikowanych jeziorach szwedzkich zawartość fosforanów jest niska. Przypuszczalną przyczyną tego jest związanie jonów fosforanowych z glinem i kumulacja ich w osadach. Z drugiej zaś strony kwaśne jeziora mogą mieć względnie wysoką zawartość azotanów. Wynika to z deficytu fosforu i związaną z tym niemożliwością wykorzystania azotanów przez rośliny w procesach metabolicznych. Niektóre inne pierwiastki, np. selen, który jest bardzo ważny w procesach wzrostowych wielu roślin wodnych, w kwaśnych jeziorach występuje w ilościach niewystarczających, ponieważ zawartość selenu zależna jest od pH wody.
Biologiczna reakcja na procesy acydyfikacyjne zachodzące w zbiornikach wodnych jest dość dobrze rozpoznana i związane z tym zmiany odnoszą się do wszystkich grup roślin i zwierząt wodnych. Glony, tak fitoplankton, jak i peryfiton, charakteryzują się zmniejszoną liczebnością gatunków, gdy pH spada poniżej 6. W silnie zakwaszonych jeziorach liczba gatunków glonów planktonowych wynosi zwykle 5-10, gdy w porównaniu z jeziorami niezakwaszonymi jest ich 30-80. Zaledwie kilka gatunków spośród zielenic nitkowatych jest zdolnych do masowej reprodukcji w takich ekstremalnych warunkach. Dobrze rozwijają się w wodzie kwaśnej torfowce (Sphagnum) i unikają pH powyżej 6. Rośliny lobeliowe (Lobelia, Isotes, Litorella) mogą tworzyć zwarte kobierce na dnie płytkich zakwaszonych jezior, jednak są one narażone na obrastanie przez torfowce i glony z zielenic nitkowatych. Trzcina (Phragmites communis), rdestnice i in. usuwają się lub znikają całkowicie. Rośliny o liściach pływających są lepiej przystosowane, ponieważ ich grube korzenie są osadzone głęboko w podłożu - korzystają z substancji odżywczych z głębszych pokładów, gdzie pH jest jeszcze wyższe.
Zwierzęta planktonowe, takie jak skorupiaki i wrotki, ubożeją w podobny sposób jak fitoplankton. W silnie zakwaszonych, przezroczystych jeziorach występują na ogół pojedyncze gatunki dominujące, zwykle spośród widłonogów (Copepoda) lub wioślarek (Cladocera). Owady wodne i ich larwy również wykazują spadek liczby gatunków w wodach zakwaszonych. Podobnie reaguje zoobentos: pijawki, skorupiaki, ślimaki i małże. Jednym z wczesnych sygnałów procesu acydyfikacji jest pojawienie się zwierząt z bardzo cienkimi pancerzami i muszlami. W związku z brakiem w kwaśnych wodach wielu gatunków ryb i innych drapieżców żerujących na larwach Chironomidae, ilość tych ostatnich wzrasta. Ryby mogą dłużej przeżyć w wodach brunatnych bogatych w humus, niż w jeziorach czystych, zakwaszonych. Przyczyna tego tkwi w możliwości fiksacji glinu przez substancje humusowe, co ogranicza toksyczne jego oddziaływanie. Żaby i ropuchy wykazują zaburzenia w rozmnażaniu w wodach kwaśnych. Nie jest jeszcze dostatecznie jasne, jaki jest bezpośredni i pośredni wpływ zakwaszenia wód na metabolizm ptaków.
Najbardziej narażone na procesy acydyfikacji są zbiorniki nieduże, wykształcone na skałach twardych, granitowych. Jak już wspomniano skutki zakwaszania są wolniejsze i mniej drastyczne, gdy w rejonie zlewni i w samym zbiorniku znajdują się pokłady węglanu wapnia. Stąd też jedną z aktualnie stosowanych metod walki z acydyfikacją wód powierzchniowych jest ich wapniowanie. Stosuje się różne metody wzbogacania wód naturalnych w wapń; dają one co prawda pozytywne efekty, są jednak bardzo kosztowne. Na podstawie szwedzkich doświadczeń uważa się, że wapniowanie jest jedynie zabiegiem profilaktycznym. W celu ograniczenia acydyfikacji trzeba wyeliminować jej przyczyny, czyli usunąć źródła emisji dwutlenku siarki, poprzez zastosowanie odpowiednich technologii w produkcji przemysłowej.
Literatura uzupełniająca
Chojnacki J.C., Podstawy ekologii wód. Akademia Rolnicza w Szczecinie, Szczecin, 1998
Demel K., Życie morza. Wydawnictwo Morskie, Gdańsk, 1974
Dussart B., Limnologia. Gauthier-Willars, Paris, 1966
Hutchinson G.E. A treatise on limnology. Wiley, New York, t.1:1957; t.2:1967
Kadłubowska J.Z., Zarys algologii. PWN, Warszawa, 1975
Kajak Z., Hydrobiologia – Limnologia, Ekosystemy wód śródlądowych. PWN, W-wa, 1998
Konstantinov A.S., Obszczaja gidrobiołogija. Wysszaja Szkoła, Moskva, 1967
Lityński A., Hydrobiologia ogólna, PWN, Warszawa, 1952
Mikulski J.S., Biologia wód śródlądowych. PWN, Warszawa, 1974
Podbielkowski Z., Tomaszewicz H., Zarys bydrobotaniki. PWN, Warszawa, 1982
Rheinheimer G., Mikrobiologia wód. PWRiL, Warszawa, 1979
Ruttner F., Grundriss der Limnologie, Gruyter, Berlin, 1962
Thorson G., Życie w morzu. Wydawnictwo Morskie, Gdańsk, 1976
Thurman H.V., Zarys oceanologii. Wydawnictwo Morskie, Gdańsk, 1982
Turoboyski L., Hydrobiologia techniczna, PWN, Warszawa, 1979
Starmach K., Metody badania planktonu. PWRiL, Warszawa, 1955
Starmach K., Wody śródlądowe. Uniwersytet Jagielloński, Kraków, 1973
Starmach K., Wróbel S., Pasternak K., Hydrobiologia. PWN, Warszawa 1976
Wolnomiejski N., Pawlikowski T., Zarys ekologii i ochrony mórz. Uniw. MK, Toruń, 2006